

Structural and kinetic analysis of protein-aggregate strains in vivo using binary epitope mapping
- PMID: 25802384
- PMCID: PMC4394267
- DOI: 10.1073/pnas.1419228112
Structural and kinetic analysis of protein-aggregate strains in vivo using binary epitope mapping
Abstract
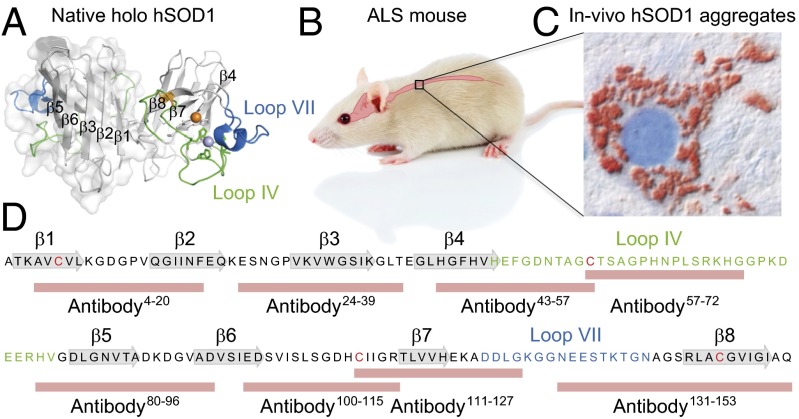
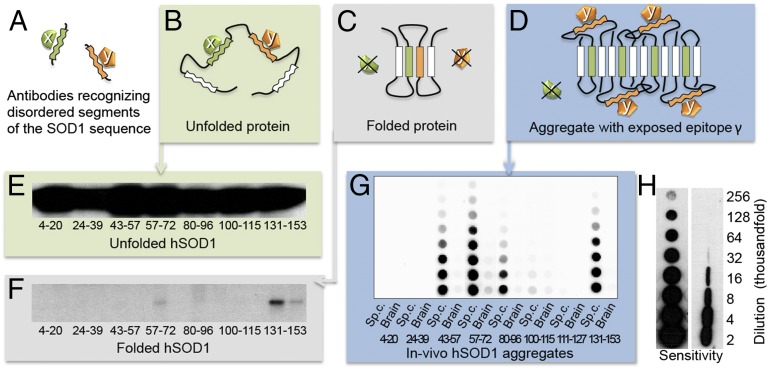
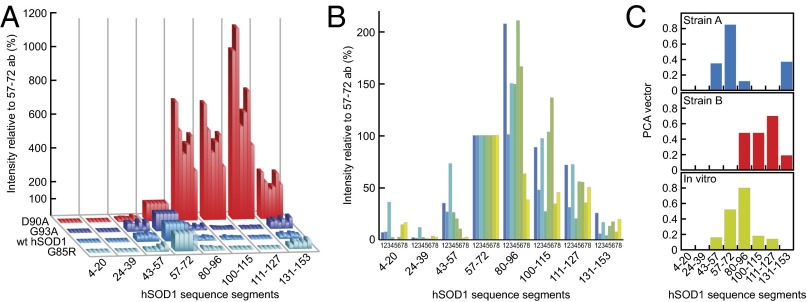
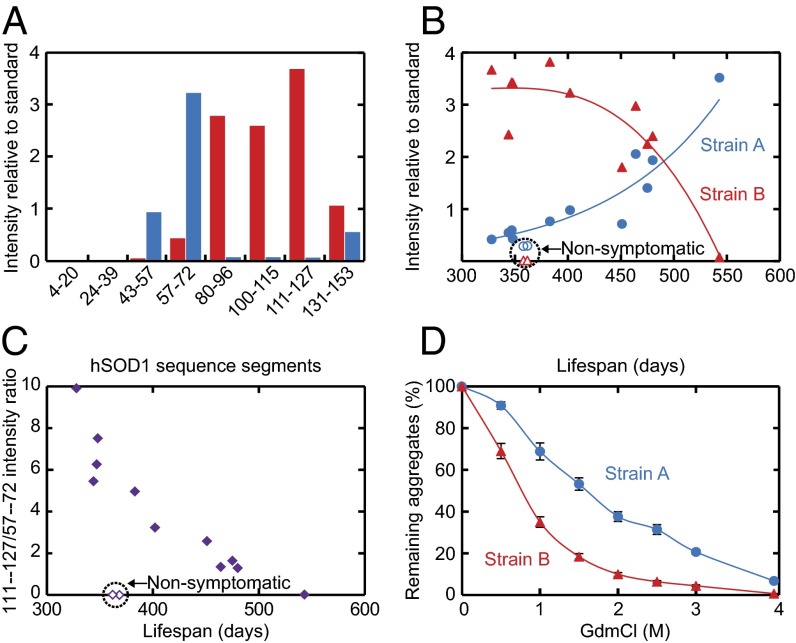
Despite considerable progress in uncovering the molecular details of protein aggregation in vitro, the cause and mechanism of protein-aggregation disease remain poorly understood. One reason is that the amount of pathological aggregates in neural tissue is exceedingly low, precluding examination by conventional approaches. We present here a method for determination of the structure and quantity of aggregates in small tissue samples, circumventing the above problem. The method is based on binary epitope mapping using anti-peptide antibodies. We assessed the usefulness and versatility of the method in mice modeling the neurodegenerative disease amyotrophic lateral sclerosis, which accumulate intracellular aggregates of superoxide dismutase-1. Two strains of aggregates were identified with different structural architectures, molecular properties, and growth kinetics. Both were different from superoxide dismutase-1 aggregates generated in vitro under a variety of conditions. The strains, which seem kinetically under fragmentation control, are associated with different disease progressions, complying with and adding detail to the growing evidence that seeding, infectivity, and strain dependence are unifying principles of neurodegenerative disease.
Keywords: amyotrophic lateral sclerosis; neurodegeneration; protein aggregation; strain; transgenic mice.
Conflict of interest statement
Conflict of interest statement: S.L.M., P.M.A., T.B., and M.O. have filed a patent partially related to concepts presented in this paper.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
