

The nicotinic receptor Alpha7 impacts the mouse lung response to LPS through multiple mechanisms
- PMID: 25803612
- PMCID: PMC4372581
- DOI: 10.1371/journal.pone.0121128
The nicotinic receptor Alpha7 impacts the mouse lung response to LPS through multiple mechanisms
Abstract
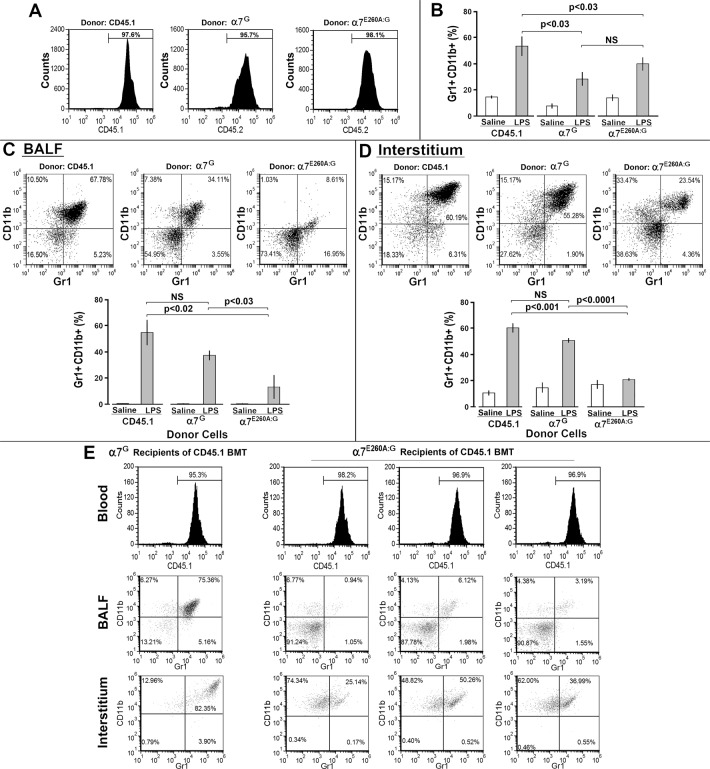
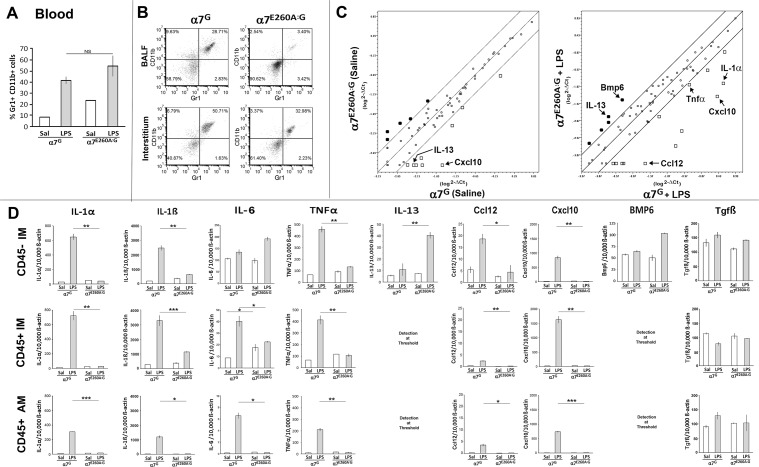
The nicotinic acetylcholine receptor alpha7 (α7) is expressed by neuronal and non-neuronal cells throughout the body. We examined the mechanisms of the lung inflammatory response to intranasal (i.n.) lipopolysaccharide (LPS) regulated by α7. This was done in mice using homologous recombination to introduce a point mutation in the α7 receptor that replaces the glutamate residue 260 that lines the pore with alanine (α7E260A), which has been implicated in controlling the exceptional calcium ion conductance of this receptor. The α7E260A mice exhibit normal inflammatory cell recruitment to the blood in response to i.n. LPS administration. This differs from the α7knock-out (α7KO) in which upstream signaling to initiate the recruitment to the blood following i.n. LPS is significantly impaired. While hematopoietic cells are recruited to the bloodstream in the α7E260A mouse, they fail to be recruited efficiently into both the interstitium and alveolar spaces of the lung. Bone marrow reconstitution experiments demonstrate that the responsiveness of both CD45+ and CD45- cells of the α7E260A mouse are impaired. The expression of several pro-inflammatory cytokine and chemokine RNAs including TNFα, IL-1α, Ccl2 and Cxcl10 are decreased in the α7E260A mouse. However, there is a substantial increase in IL-13 expression by CD45- lung interstitial cells in the α7E260A mouse. Our results support the conclusion that α7 functional pleiotropy contributes to modulating the tissue response to an inflammatory insult through impacting upon a variety of mechanisms reflecting the individual cell composition of the lung.
Conflict of interest statement
Figures
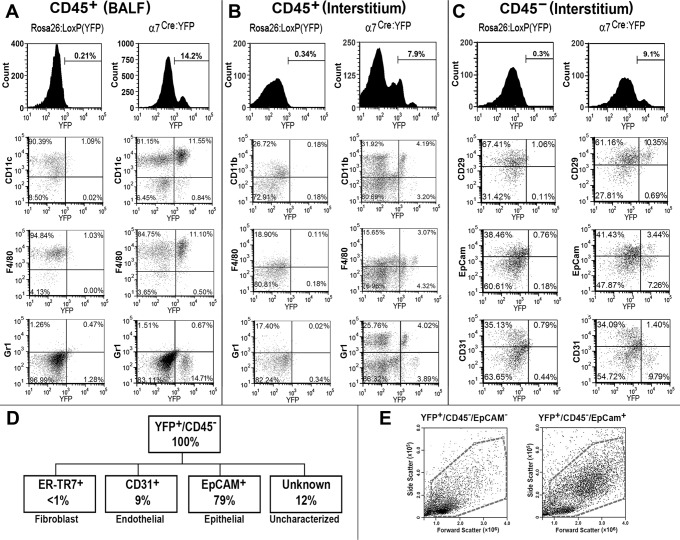
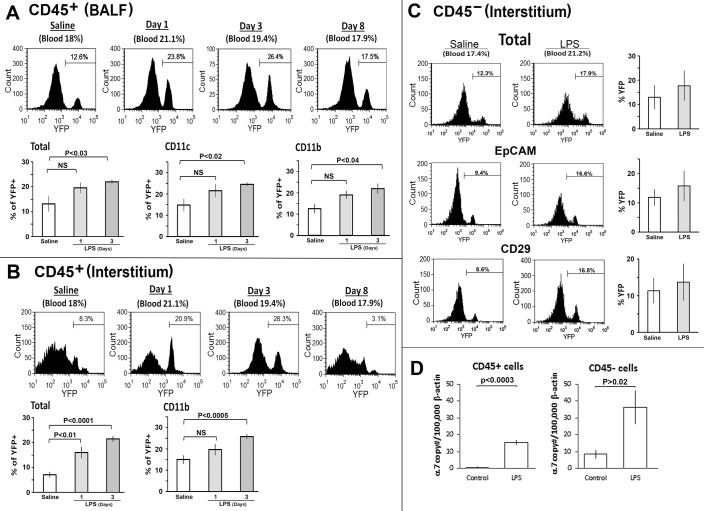
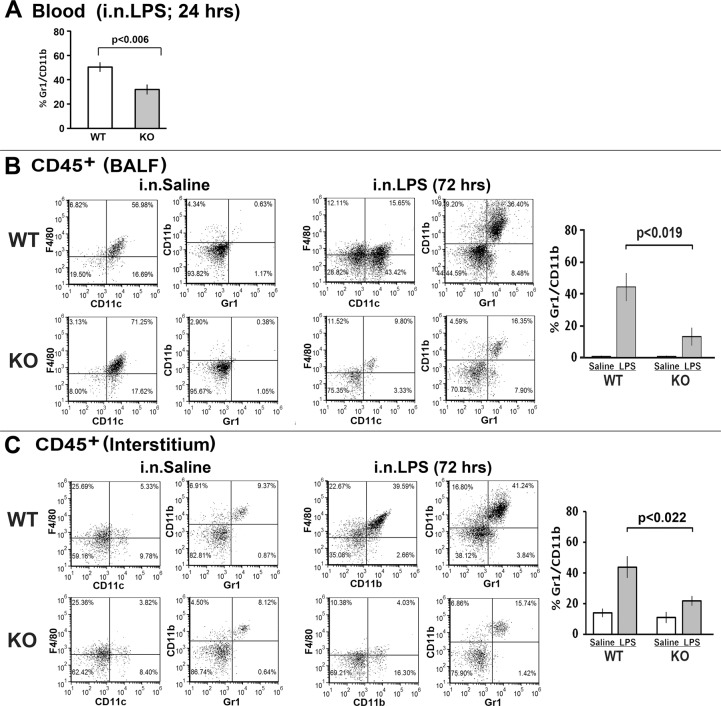
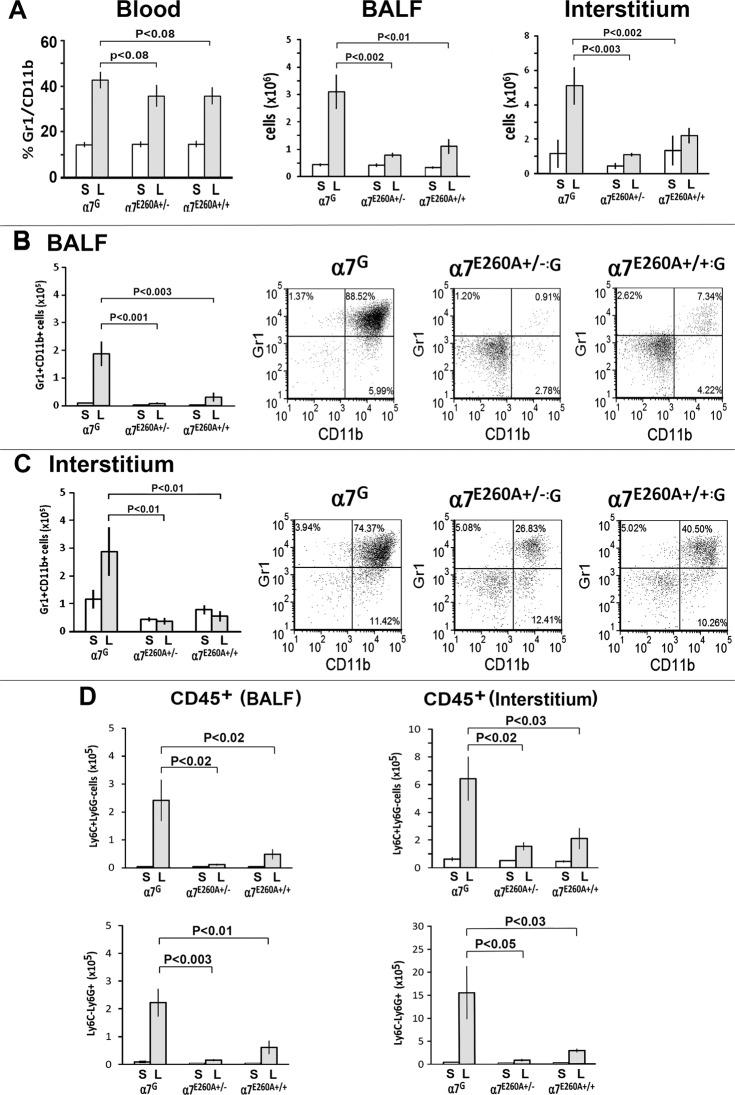
References
-
- Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, et al. (2003) Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 421: 384–388. - PubMed
-
- Pavlov VA, Tracey KJ (2005) The cholinergic anti-inflammatory pathway. Brain Behav Immun 19: 493–499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
