

AMBRA1 and BECLIN 1 interplay in the crosstalk between autophagy and cell proliferation
- PMID: 25803737
- PMCID: PMC4615147
- DOI: 10.1080/15384101.2015.1021526
AMBRA1 and BECLIN 1 interplay in the crosstalk between autophagy and cell proliferation
Abstract
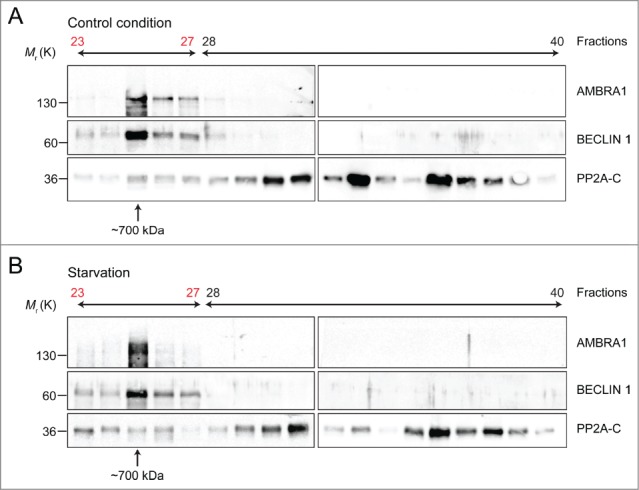
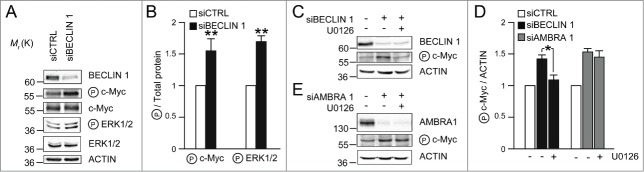
Autophagy-promoting proteins and stimuli are often associated with inhibition of cell proliferation; in this context, we recently described a key role for the pro-autophagic protein AMBRA1. Indeed, AMBRA1, through its direct interaction with the protein phosphatase PP2A, tightly regulates the stability of the oncoprotein and pro-mitotic factor c-Myc. Moreover, the AMBRA1-mediated regulation of c-Myc affects both cell proliferation rate and tumorigenesis. Interestingly, AMBRA1/PP2A activity is under the control of the master regulator of autophagy and cell growth, the protein kinase mTOR. Besides the mechanistic details of this regulation pathway which we dissected previously, any possible interplay(s) between AMBRA1 and its interactor BECLIN 1 was not investigated in this scenario. Here we show that both AMBRA1 and BECLIN 1 affect c-Myc regulation, but through two different pathways. Nevertheless, these two pro-autophagic proteins are, together with PP2A, in the same macromolecular complex, whose functional significance of which will be addressed in future studies.
Keywords: EGFR degradation; PP2A; c-Myc; cancer; cell cycle.
Figures
Comment in
-
AMBRA1: When autophagy meets cell proliferation.Autophagy. 2015;11(9):1705-7. doi: 10.1080/15548627.2015.1053681. Autophagy. 2015. PMID: 26101901 Free PMC article.
References
-
- Boya P, Reggiori F, Codogno P. Emerging regulation and functions of autophagy. Nat Cell Biol 2013; 15:713–20; PMID:23817233; http://dx.doi.org/ 10.1038/ncb2788 - DOI - PMC - PubMed
-
- Abada A, Elazar Z. Getting ready for building: signaling and autophagosome biogenesis. EMBO Rep 2014; 15:839–52; PMID:25027988; http://dx.doi.org/ 10.15252/embr.201439076 - DOI - PMC - PubMed
-
- Kroemer G, Marino G, Levine B. Autophagy and the integrated stress response. Mol Cell 2010; 40:280–93; PMID:20965422; http://dx.doi.org/ 10.1016/j.molcel.2010.09.023 - DOI - PMC - PubMed
-
- Marino G, Niso-Santano M, Baehrecke EH, Kroemer G. Self-consumption: the interplay of autophagy and apoptosis. Nat Rev Mol Cell Biol 2014; 15:81–94; PMID:24401948; http://dx.doi.org/ 10.1038/nrm3735 - DOI - PMC - PubMed
-
- Cianfanelli V, Cecconi F. Autophagy-dependent NFkappaB regulation. Cell Cycle 2012; 11:436–7; PMID:22262191; http://dx.doi.org/ 10.4161/cc.11.3.19224 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous