

Opposite phenotypes of muscle strength and locomotor function in mouse models of partial trisomy and monosomy 21 for the proximal Hspa13-App region
- PMID: 25803843
- PMCID: PMC4372517
- DOI: 10.1371/journal.pgen.1005062
Opposite phenotypes of muscle strength and locomotor function in mouse models of partial trisomy and monosomy 21 for the proximal Hspa13-App region
Abstract
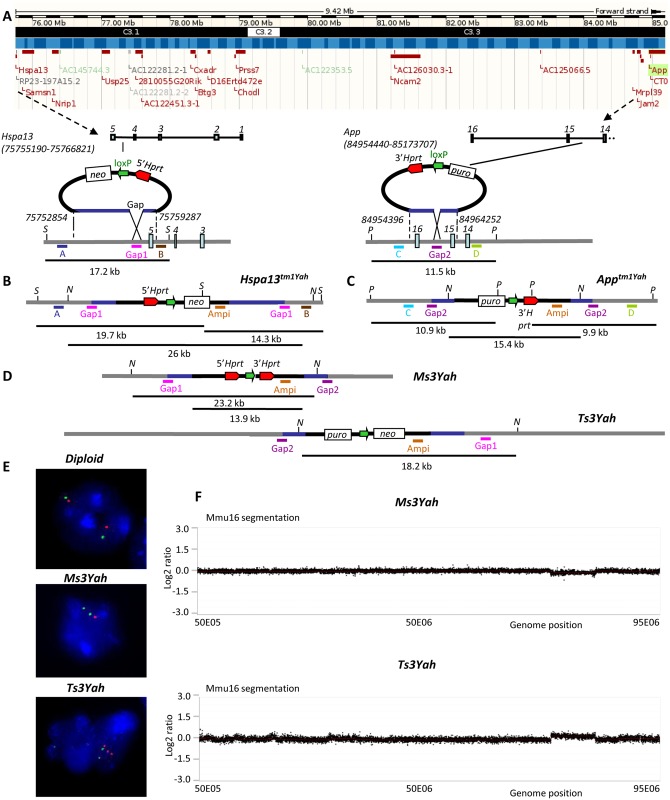
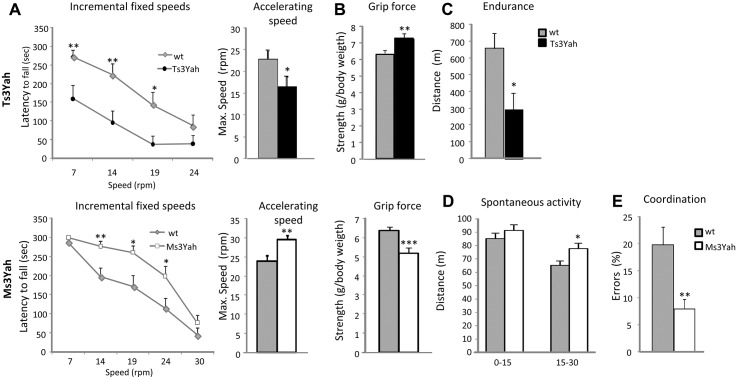
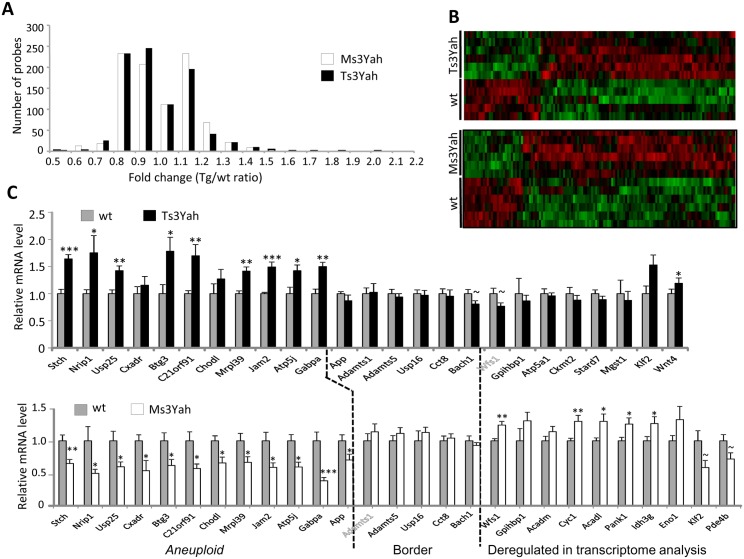
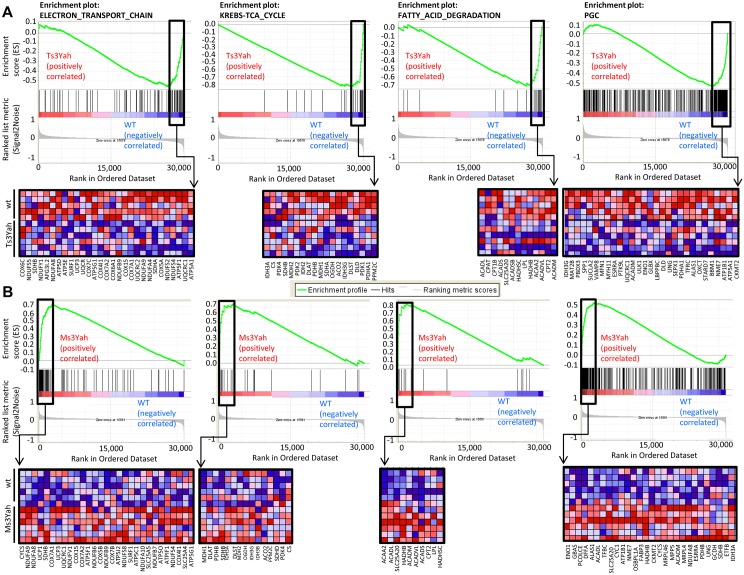
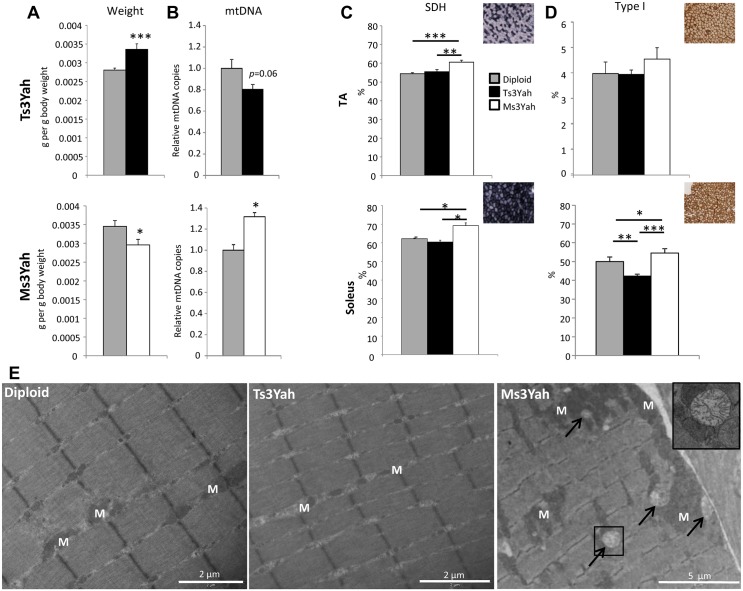
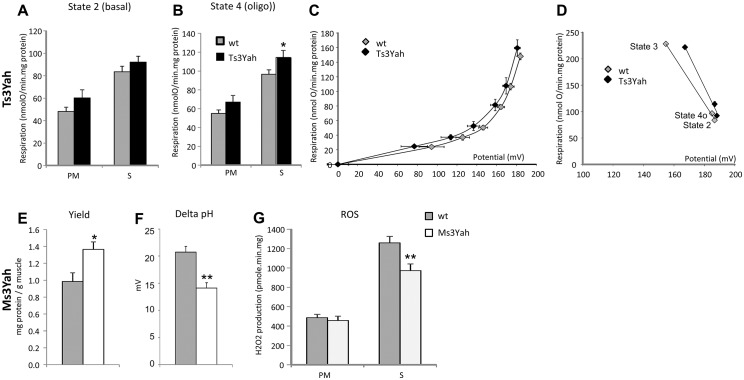
The trisomy of human chromosome 21 (Hsa21), which causes Down syndrome (DS), is the most common viable human aneuploidy. In contrast to trisomy, the complete monosomy (M21) of Hsa21 is lethal, and only partial monosomy or mosaic monosomy of Hsa21 is seen. Both conditions lead to variable physiological abnormalities with constant intellectual disability, locomotor deficits, and altered muscle tone. To search for dosage-sensitive genes involved in DS and M21 phenotypes, we created two new mouse models: the Ts3Yah carrying a tandem duplication and the Ms3Yah carrying a deletion of the Hspa13-App interval syntenic with 21q11.2-q21.3. Here we report that the trisomy and the monosomy of this region alter locomotion, muscle strength, mass, and energetic balance. The expression profiling of skeletal muscles revealed global changes in the regulation of genes implicated in energetic metabolism, mitochondrial activity, and biogenesis. These genes are downregulated in Ts3Yah mice and upregulated in Ms3Yah mice. The shift in skeletal muscle metabolism correlates with a change in mitochondrial proliferation without an alteration in the respiratory function. However, the reactive oxygen species (ROS) production from mitochondrial complex I decreased in Ms3Yah mice, while the membrane permeability of Ts3Yah mitochondria slightly increased. Thus, we demonstrated how the Hspa13-App interval controls metabolic and mitochondrial phenotypes in muscles certainly as a consequence of change in dose of Gabpa, Nrip1, and Atp5j. Our results indicate that the copy number variation in the Hspa13-App region has a peripheral impact on locomotor activity by altering muscle function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Manolakos E, Peitsidis P, Eleftheriades M, Dedoulis E, Ziegler M, Orru S, et al. Prenatal detection of full monosomy 21 in a fetus with increased nuchal translucency: Molecular cytogenetic analysis and review of the literature. Journal of Obstetrics and Gynaecology Research. 2010;36(2):435–40. 10.1111/j.1447-0756.2009.01140.x - DOI - PubMed
-
- Mori MA, Lapunzina P, Delicado A, Nunez G, Rodriguez JI, de Torres ML, et al. A prenatally diagnosed patient with full monosomy 21: Ultrasound, cytogenetic, clinical, molecular, and necropsy findings. American Journal of Medical Genetics Part A. 2004;127A(1):69–73. 10.1002/ajmg.a.20622 - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
