

Cathepsin B promotes colorectal tumorigenesis, cell invasion, and metastasis
- PMID: 25808857
- PMCID: PMC4832390
- DOI: 10.1002/mc.22312
Cathepsin B promotes colorectal tumorigenesis, cell invasion, and metastasis
Abstract
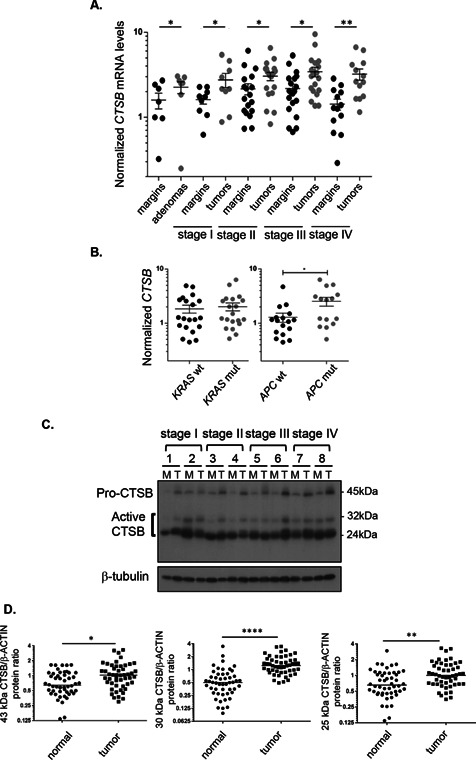
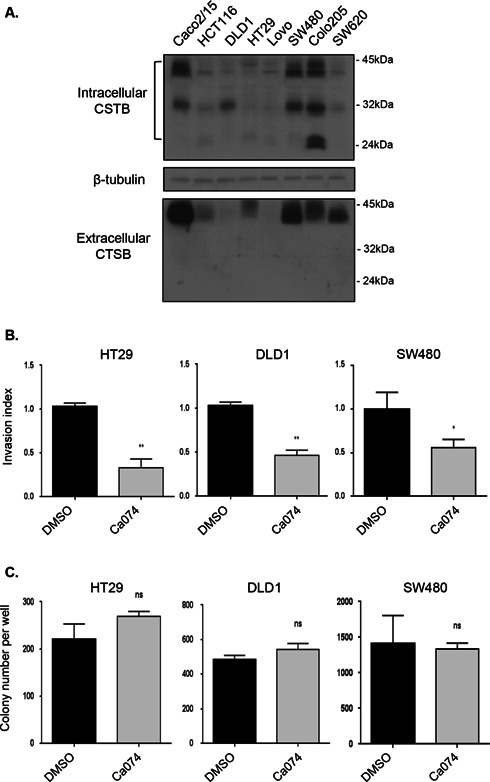
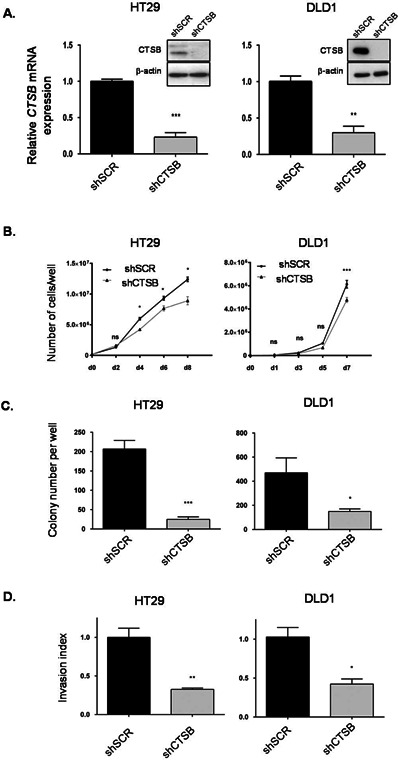
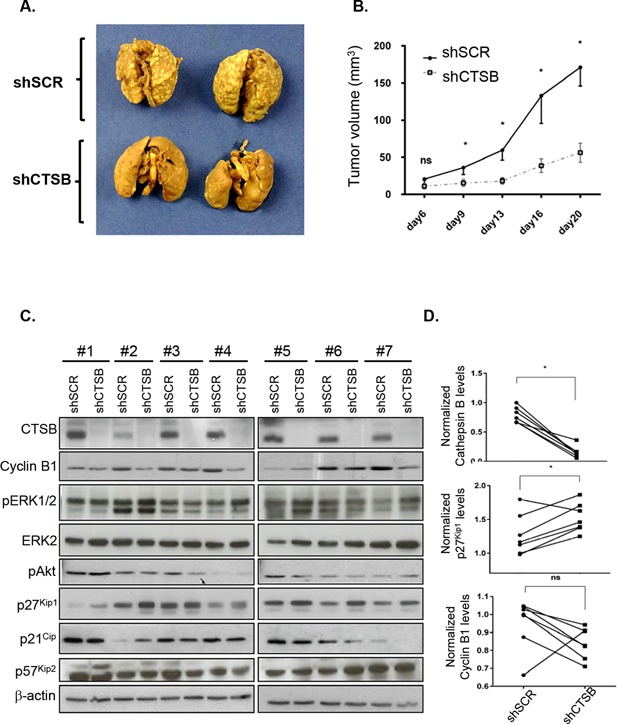
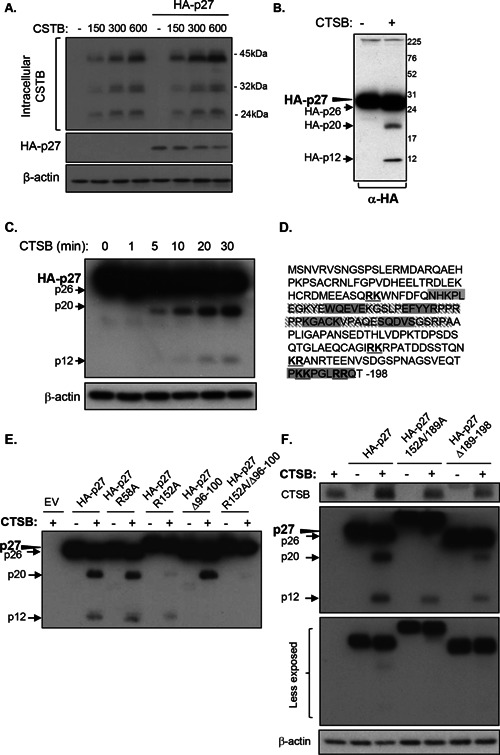
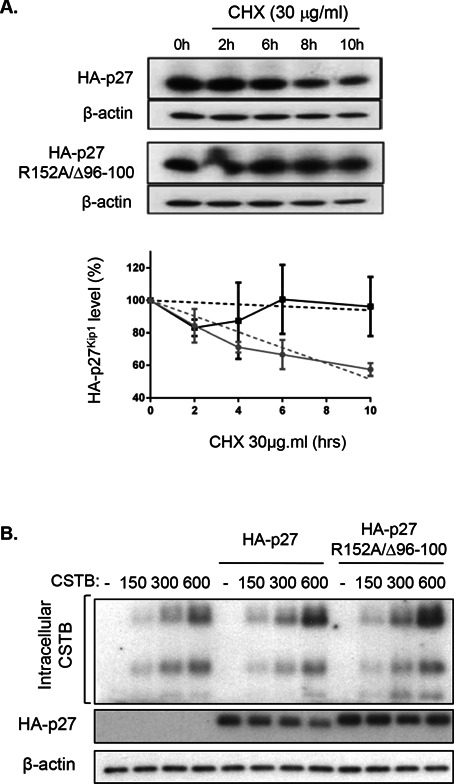
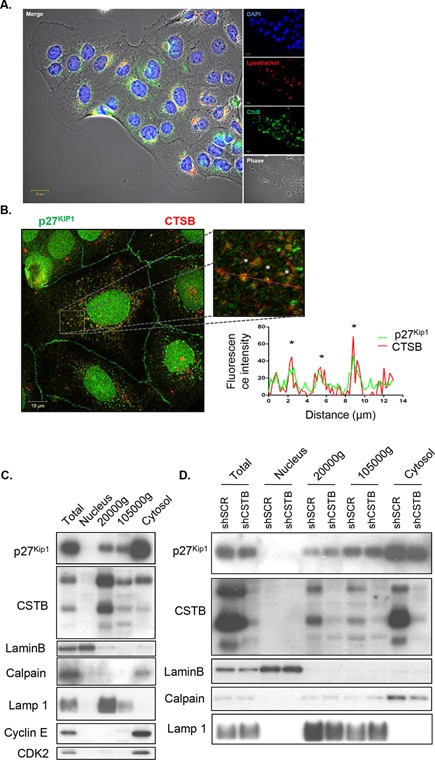
Cathepsin B is a cysteine proteinase that primarily functions as an endopeptidase within endolysosomal compartments in normal cells. However, during tumoral expansion, the regulation of cathepsin B can be altered at multiple levels, thereby resulting in its overexpression and export outside of the cell. This may suggest a possible role of cathepsin B in alterations leading to cancer progression. The aim of this study was to determine the contribution of intracellular and extracellular cathepsin B in growth, tumorigenesis, and invasion of colorectal cancer (CRC) cells. Results show that mRNA and activated levels of cathepsin B were both increased in human adenomas and in CRCs of all stages. Treatment of CRC cells with the highly selective and non-permeant cathepsin B inhibitor Ca074 revealed that extracellular cathepsin B actively contributed to the invasiveness of human CRC cells while not essential for their growth in soft agar. Cathepsin B silencing by RNAi in human CRC cells inhibited their growth in soft agar, as well as their invasion capacity, tumoral expansion, and metastatic spread in immunodeficient mice. Higher levels of the cell cycle inhibitor p27(Kip1) were observed in cathepsin B-deficient tumors as well as an increase in cyclin B1. Finally, cathepsin B colocalized with p27(Kip1) within the lysosomes and efficiently degraded the inhibitor. In conclusion, the present data demonstrate that cathepsin B is a significant factor in colorectal tumor development, invasion, and metastatic spreading and may, therefore, represent a potential pharmacological target for colorectal tumor therapy.
Keywords: cathepsin B; colorectal cancers; intestinal tumorigenesis; invasion; p27Kip1.
© 2015 The Authors. Molecular Carcinogenesis Published by Wiley Periodicals, Inc.
Figures
Similar articles
-
microRNA-200b and microRNA-200c promote colorectal cancer cell proliferation via targeting the reversion-inducing cysteine-rich protein with Kazal motifs.RNA Biol. 2015;12(3):276-89. doi: 10.1080/15476286.2015.1017208. RNA Biol. 2015. PMID: 25826661 Free PMC article.
-
OVOL2, an Inhibitor of WNT Signaling, Reduces Invasive Activities of Human and Mouse Cancer Cells and Is Down-regulated in Human Colorectal Tumors.Gastroenterology. 2016 Mar;150(3):659-671.e16. doi: 10.1053/j.gastro.2015.11.041. Epub 2015 Nov 24. Gastroenterology. 2016. PMID: 26619963
-
Positive feedback loop of hepatoma-derived growth factor and β-catenin promotes carcinogenesis of colorectal cancer.Oncotarget. 2015 Oct 6;6(30):29357-74. doi: 10.18632/oncotarget.4982. Oncotarget. 2015. PMID: 26296979 Free PMC article.
-
Cathepsin B: multiple roles in cancer.Proteomics Clin Appl. 2014 Jun;8(5-6):427-37. doi: 10.1002/prca.201300105. Epub 2014 Mar 26. Proteomics Clin Appl. 2014. PMID: 24677670 Free PMC article. Review.
-
Cathepsin B: A sellsword of cancer progression.Cancer Lett. 2019 May 1;449:207-214. doi: 10.1016/j.canlet.2019.02.035. Epub 2019 Feb 20. Cancer Lett. 2019. PMID: 30796968 Free PMC article. Review.
Cited by
-
The Significance of Cathepsin B in Mediating Radiation Resistance in Colon Carcinoma Cell Line (Caco-2).Int J Mol Sci. 2023 Nov 9;24(22):16146. doi: 10.3390/ijms242216146. Int J Mol Sci. 2023. PMID: 38003335 Free PMC article.
-
Enzyme-Responsive Amphiphilic Peptide Nanoparticles for Biocompatible and Efficient Drug Delivery.Pharmaceutics. 2022 Jan 7;14(1):143. doi: 10.3390/pharmaceutics14010143. Pharmaceutics. 2022. PMID: 35057039 Free PMC article.
-
The Role of Cathepsin B in Pathophysiologies of Non-tumor and Tumor tissues: A Systematic Review.J Cancer. 2023 Jul 24;14(12):2344-2358. doi: 10.7150/jca.86531. eCollection 2023. J Cancer. 2023. PMID: 37576397 Free PMC article. Review.
-
CTSG Suppresses Colorectal Cancer Progression through Negative Regulation of Akt/mTOR/Bcl2 Signaling Pathway.Int J Biol Sci. 2023 Apr 17;19(7):2220-2233. doi: 10.7150/ijbs.82000. eCollection 2023. Int J Biol Sci. 2023. PMID: 37151875 Free PMC article.
-
Over-expression of cathepsin B in hepatocellular carcinomas predicts poor prognosis of HCC patients.Mol Cancer. 2016 Feb 20;15:17. doi: 10.1186/s12943-016-0503-9. Mol Cancer. 2016. PMID: 26896959 Free PMC article.
References
-
- Fearon ER, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell 1990; 61:759–767. - PubMed
-
- Yan S, Sloane BF. Molecular regulation of human cathepsin B: Implication in pathologies. Biol Chem 2003; 384:845–854. - PubMed
-
- Hazen LG, Bleeker FE, Lauritzen B, et al. Comparative localization of cathepsin B protein and activity in colorectal cancer. J Histochem Cytochem 2000; 48:1421–1430. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
