

LKB1/Mo25/STRAD uniquely impacts sarcomeric contractile function and posttranslational modification
- PMID: 25809261
- PMCID: PMC4375436
- DOI: 10.1016/j.bpj.2015.02.012
LKB1/Mo25/STRAD uniquely impacts sarcomeric contractile function and posttranslational modification
Abstract
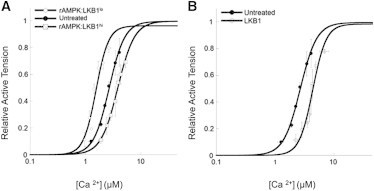
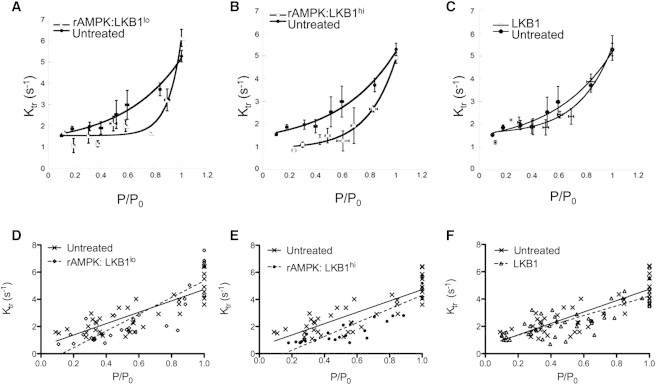
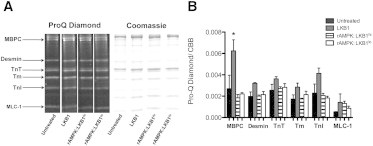
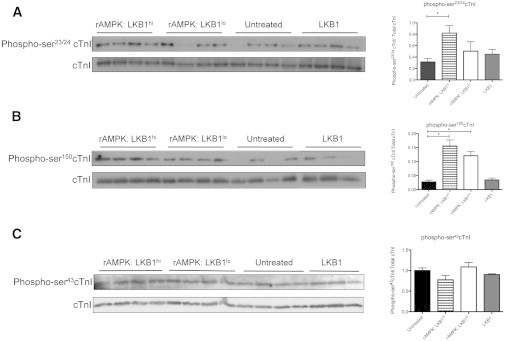
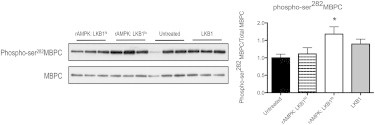
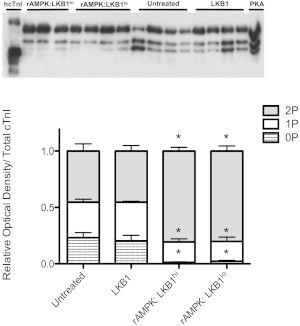
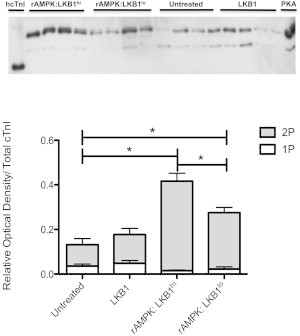
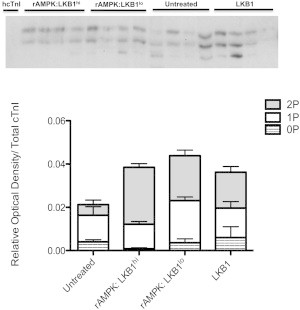
The myocardium undergoes extensive metabolic and energetic remodeling during the progression of cardiac disease. Central to remodeling are changes in the adenine nucleotide pool. Fluctuations in these pools can activate AMP-activated protein kinase (AMPK), the central regulator of cellular energetics. Binding of AMP to AMPK not only allosterically activates AMPK but also promotes phosphorylation of AMPK by an upstream kinase complex, LKB1/Mo25/STRAD (liver kinase B 1, mouse protein 25, STE-related adaptor protein). AMPK phosphorylation by the LKB1 complex results in a substantial increase in AMPK activity. Molecular targeting by the LKB1 complex depends on subcellular localization and transcriptional expression. Yet, little is known about the ability of the LKB1 complex to modulate targeting of AMPK after activation. Accordingly, we hypothesized that differing stoichiometric ratios of LKB1 activator complex to AMPK would uniquely impact myofilament function. Demembranated rat cardiac trabeculae were incubated with varying ratios of the LKB1 complex to AMPK or the LKB1 complex alone. After incubation, we measured the Ca(2+) sensitivity of tension, rate constant for tension redevelopment, maximum tension generation, length-dependent activation, cooperativity, and sarcomeric protein phosphorylation status. We found that the Ca(2+) sensitivity of tension and cross-bridge dynamics were dependent on the LKB1 complex/AMPK ratio. We also found that the LKB1 complex desensitizes and suppresses myofilament function independently of AMPK. A phospho-proteomic analysis of myofilament proteins revealed site-specific changes in cardiac Troponin I (cTnI) phosphorylation, as well as a unique distribution of cTnI phosphospecies that were dependent on the LKB1 complex/ AMPK ratio. Fibers treated with the LKB1 complex alone did not alter cTnI phosphorylation or phosphospecies distribution. However, LKB1 complex treatment independent of AMPK increased phosphorylation of myosin-binding protein C. Therefore, we conclude that the LKB1/AMPK signaling axis is able to alter muscle function through multiple mechanisms.
Copyright © 2015 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Nascimben L., Ingwall J.S., Allen P.D. Creatine kinase system in failing and nonfailing human myocardium. Circulation. 1996;94:1894–1901. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
