

Mechanisms of epithelial wound detection
- PMID: 25813429
- PMCID: PMC4475481
- DOI: 10.1016/j.tcb.2015.02.007
Mechanisms of epithelial wound detection
Abstract
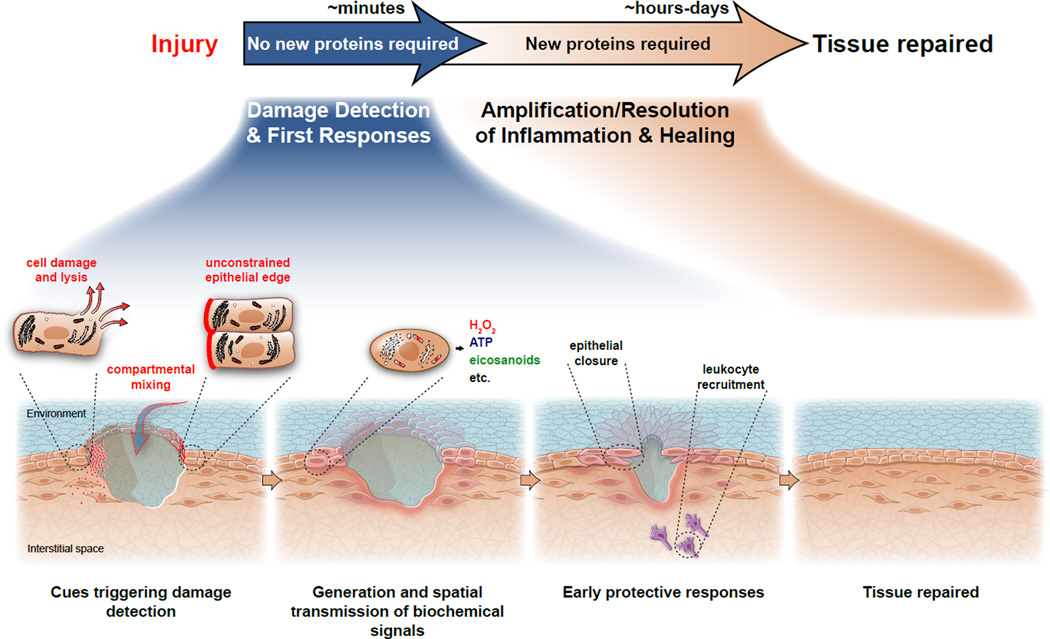
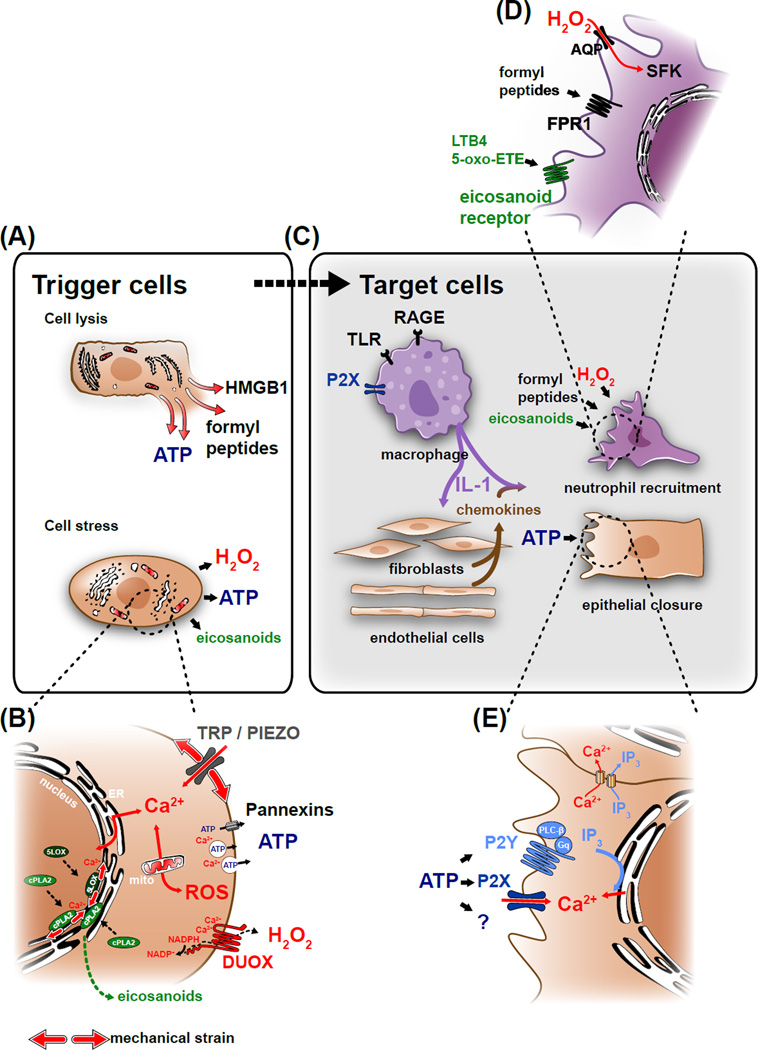
Efficient wound healing requires the coordinated responses of various cell types within an injured tissue. To react to the presence of a wound, cells have to first detect it. Judging from their initial biochemical and morphological responses, many cells including leukocytes, epithelial cells, and endothelial cells detect wounds from over hundreds of micrometers within seconds-to-minutes. Wound detection involves the conversion of an injury-induced homeostatic perturbation, such as cell lysis, an unconstrained epithelial edge, or permeability barrier breakdown, into a chemical or physical signal. The signal is spatially propagated through the tissue to synchronize protective responses of cells near the wound site and at a distance. This review summarizes the triggers and mechanisms of wound detection in animals.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Szpaderska AM, et al. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003;82:621–626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
