

Brg1 modulates enhancer activation in mesoderm lineage commitment
- PMID: 25813539
- PMCID: PMC4392595
- DOI: 10.1242/dev.109496
Brg1 modulates enhancer activation in mesoderm lineage commitment
Abstract
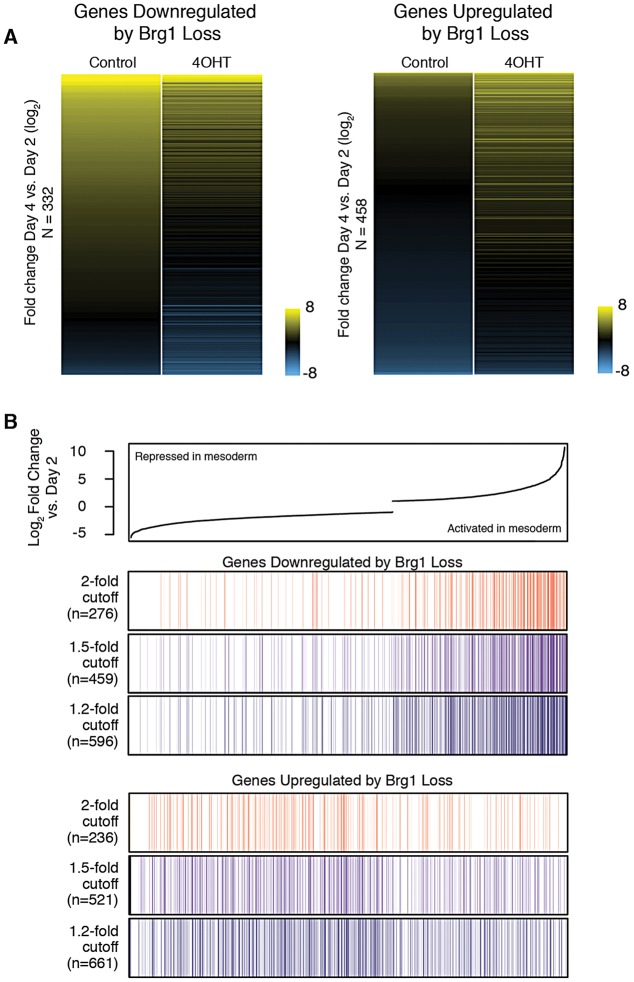
The interplay between different levels of gene regulation in modulating developmental transcriptional programs, such as histone modifications and chromatin remodeling, is not well understood. Here, we show that the chromatin remodeling factor Brg1 is required for enhancer activation in mesoderm induction. In an embryonic stem cell-based directed differentiation assay, the absence of Brg1 results in a failure of cardiomyocyte differentiation and broad deregulation of lineage-specific gene expression during mesoderm induction. We find that Brg1 co-localizes with H3K27ac at distal enhancers and is required for robust H3K27 acetylation at distal enhancers that are activated during mesoderm induction. Brg1 is also required to maintain Polycomb-mediated repression of non-mesodermal developmental regulators, suggesting cooperativity between Brg1 and Polycomb complexes. Thus, Brg1 is essential for modulating active and repressive chromatin states during mesoderm lineage commitment, in particular the activation of developmentally important enhancers. These findings demonstrate interplay between chromatin remodeling complexes and histone modifications that, together, ensure robust and broad gene regulation during crucial lineage commitment decisions.
Keywords: Chromatin; Enhancers; Gene expression; Histone modification; Mesoderm; Stem cells.
© 2015. Published by The Company of Biologists Ltd.
Figures







References
-
- Attanasio C., Nord A. S., Zhu Y., Blow M. J., Biddie S. C., Mendenhall E. M., Dixon J., Wright C., Hosseini R., Akiyama J. A. et al. (2014). Tissue-specific SMARCA4 binding at active and repressed regulatory elements during embryogenesis. Genome Res. 24, 920-929 10.1101/gr.168930.113 - DOI - PMC - PubMed
-
- Bultman S., Gebuhr T., Yee D., La Mantia C., Nicholson J., Gilliam A., Randazzo F., Metzger D., Chambon P., Crabtree G. et al. (2000). A Brg1 null mutation in the mouse reveals functional differences among mammalian SWI/SNF complexes. Mol. Cell 6, 1287-1295 10.1016/S1097-2765(00)00127-1 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
