

Nonbiased Molecular Screening Identifies Novel Molecular Regulators of Fibrogenic and Proliferative Signaling in Myxomatous Mitral Valve Disease
- PMID: 25814644
- PMCID: PMC4640679
- DOI: 10.1161/CIRCGENETICS.114.000921
Nonbiased Molecular Screening Identifies Novel Molecular Regulators of Fibrogenic and Proliferative Signaling in Myxomatous Mitral Valve Disease
Abstract
Background: Pathological processes underlying myxomatous mitral valve degeneration (MMVD) remain poorly understood. We sought to identify novel mechanisms contributing to the development of this condition.
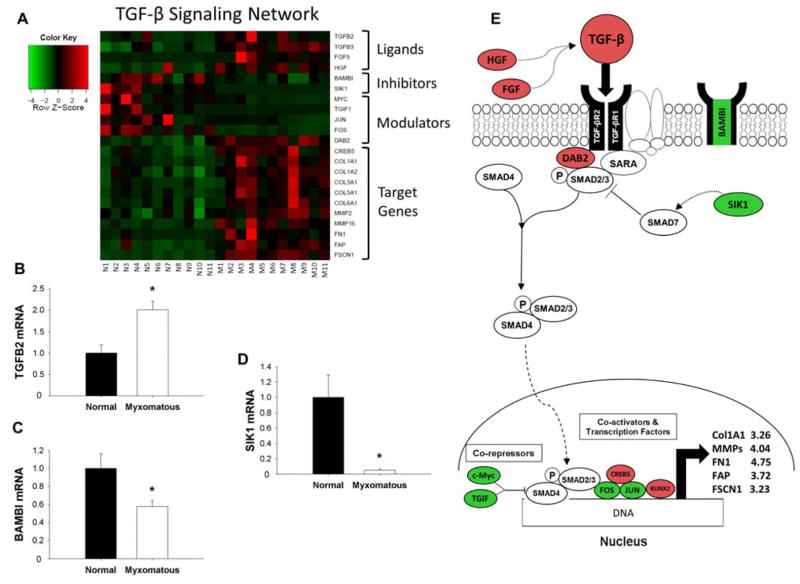
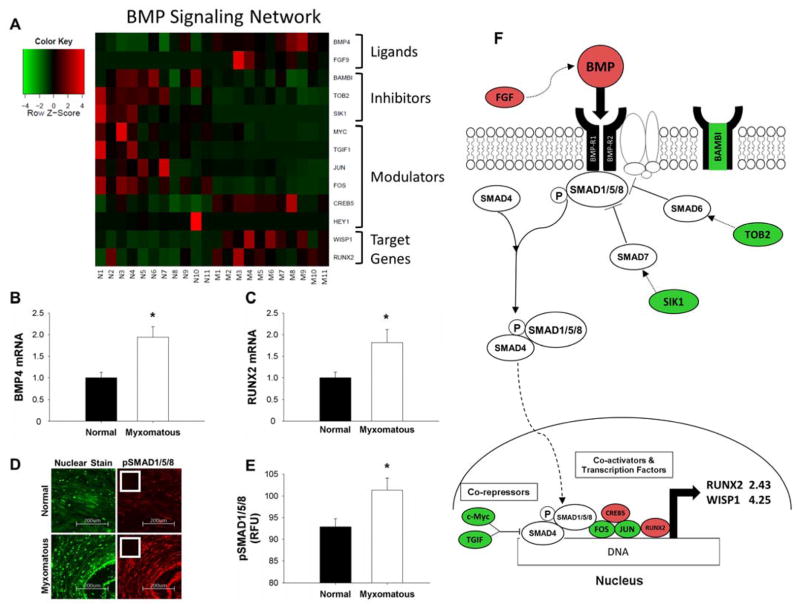
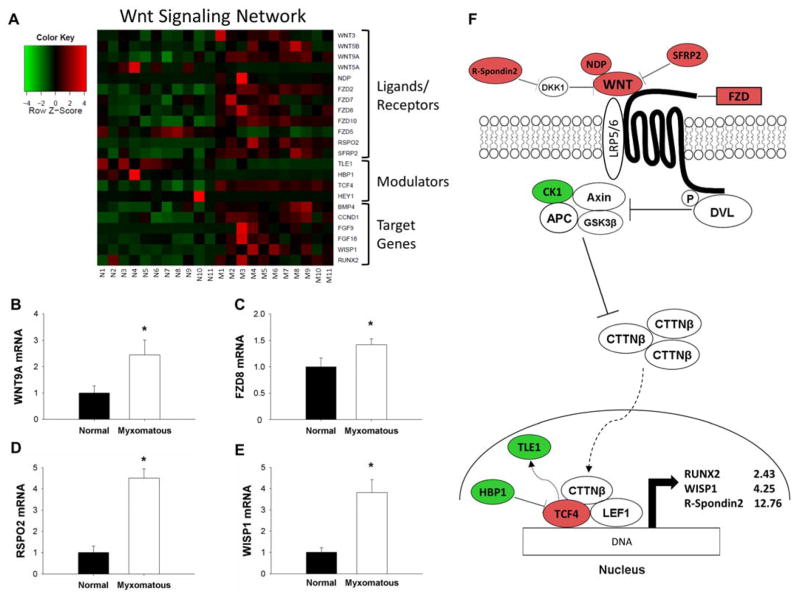
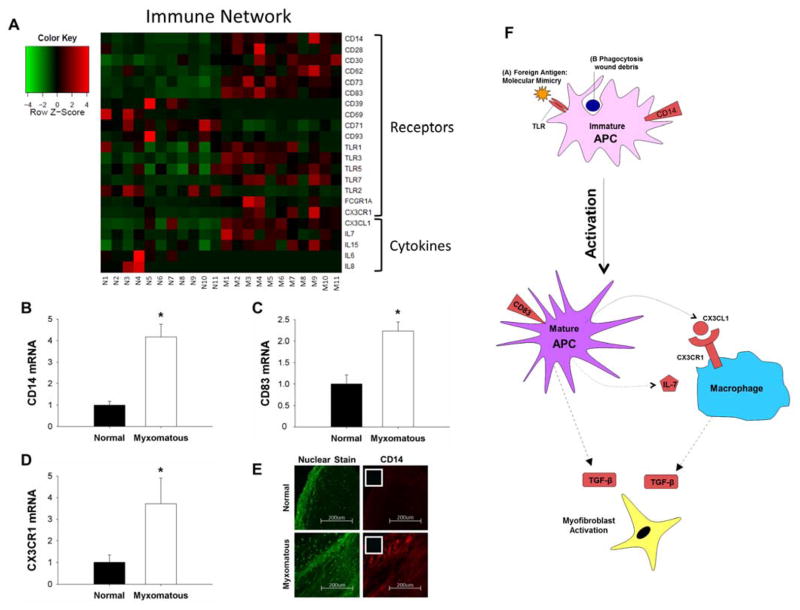
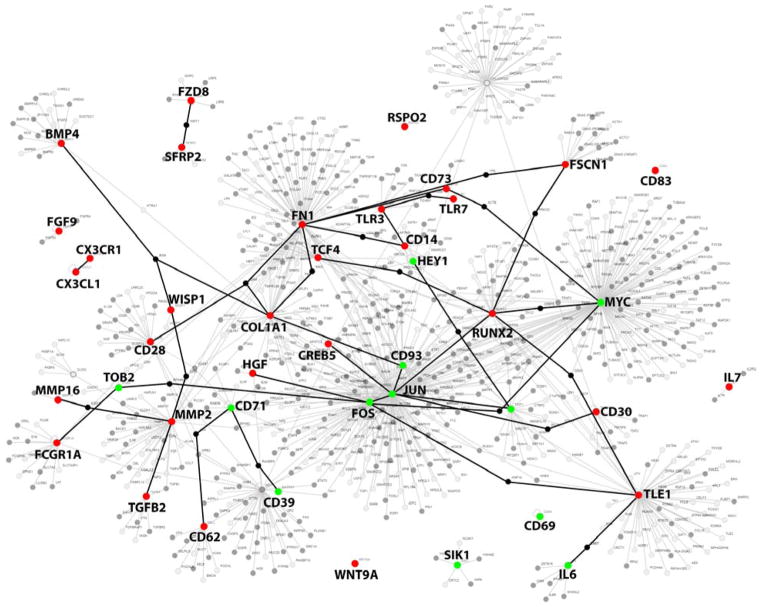
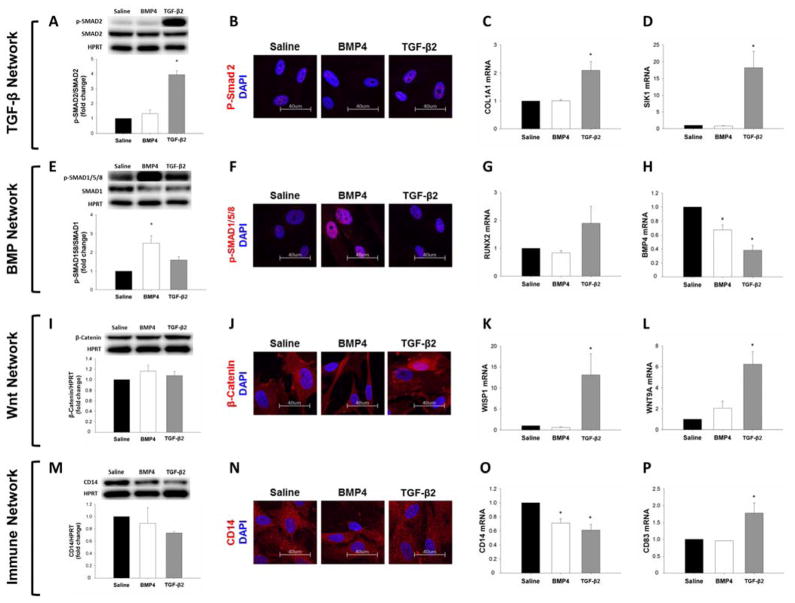
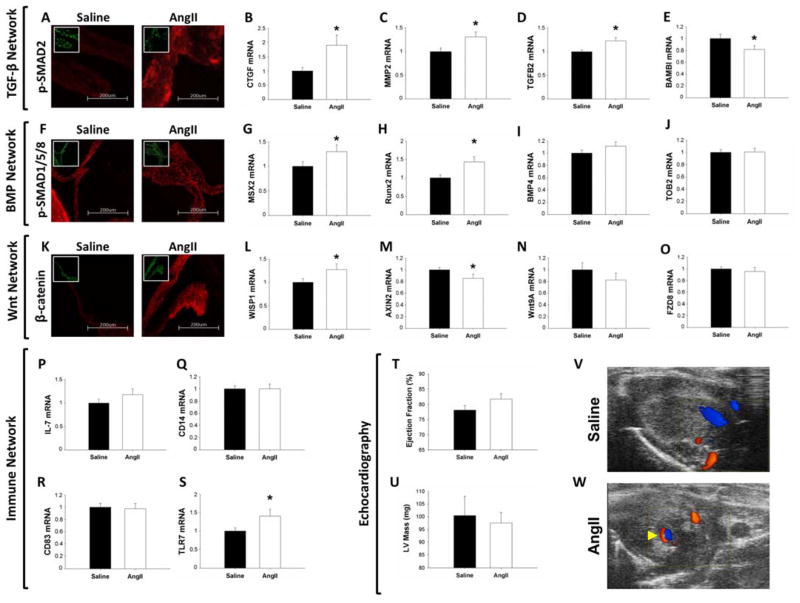
Methods and results: Microarrays were used to measure gene expression in 11 myxomatous and 11 nonmyxomatous human mitral valves. Differential gene expression (thresholds P<0.05; fold-change >1.5) and pathway activation (Ingenuity) were confirmed using quantitative reverse transcriptase polymerase chain reaction and immunohistochemistry. Contributions of bone morphogenetic protein 4 and transforming growth factor (TGF)-β2 to differential gene expression were evaluated in vitro. Contributions of angiotensin II to differential pathway activation were examined in mice in vivo. A total of 2602 genes were differentially expressed between myxomatous and nonmyxomatous valves. Canonical TGF-β signaling was increased in MMVD because of increased ligand expression and derepression of SMA mothers against decapentaplegic 2/3 signaling and was confirmed with quantitative reverse transcriptase polymerase chain reaction and immunohistochemistry. Myxomatous valves demonstrated activation of canonical bone morphogenetic protein and Wnt/β-catenin signaling and upregulation of their common target runt-related transcription factor 2. Our data set provided transcriptional and immunohistochemical evidence for activated immune cell infiltration. In vitro treatment of mitral valve interstitial cells with TGF-β2 increased β-catenin signaling at mRNA and protein levels, suggesting interactions between TGF-β2 and Wnt signaling. In vivo infusion of mice with angiotensin II recaptured several changes in signaling pathways characteristic of human MMVD.
Conclusions: These data support a new disease framework whereby activation of TGF-β2, bone morphogenetic protein 4, Wnt/β-catenin, or immune signaling plays major roles in the pathogenesis of MMVD. We propose these pathways act in a context-dependent manner to drive phenotypic changes that fundamentally differ from those observed in aortic valve disease and open novel avenues guiding future research into the pathogenesis of MMVD.
Keywords: general surgery; mitral valve; molecular biology; pathology.
© 2015 American Heart Association, Inc.
Figures
References
-
- Freed LA, Levy D, Levine RA, Larson MG, Evans JC, Fuller DL, et al. Prevalence and Clinical Outcome of Mitral Valve Prolapse. N Engl J Med. 1999;341:1–7. - PubMed
-
- Guthrie RB, Edwards JE. Pathology of the Myxomatous Mitral Valve: Nature, Secondary Changes and Complications. Minn Med. 1976;59:637–647. - PubMed
-
- Avierinos J, Gersh BJ, Melton JI, Bailey KR, Shub C, Nishimura RA, et al. Natural History of Asymptomatic Mitral Valve Prolapse in the Community. Circulation. 2002;106:1355–1361. - PubMed
-
- Suri RM, Schaff HV, Dearani Ja, Sundt TM, Daly RC, Mullany CJ, et al. Survival advantage and improved durability of mitral repair for leaflet prolapse subsets in the current era. Ann Thorac Surg. 2006;82:819–826. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
