

Promiscuous actions of small molecule inhibitors of the protein kinase D-class IIa HDAC axis in striated muscle
- PMID: 25816750
- PMCID: PMC4410688
- DOI: 10.1016/j.febslet.2015.03.017
Promiscuous actions of small molecule inhibitors of the protein kinase D-class IIa HDAC axis in striated muscle
Abstract
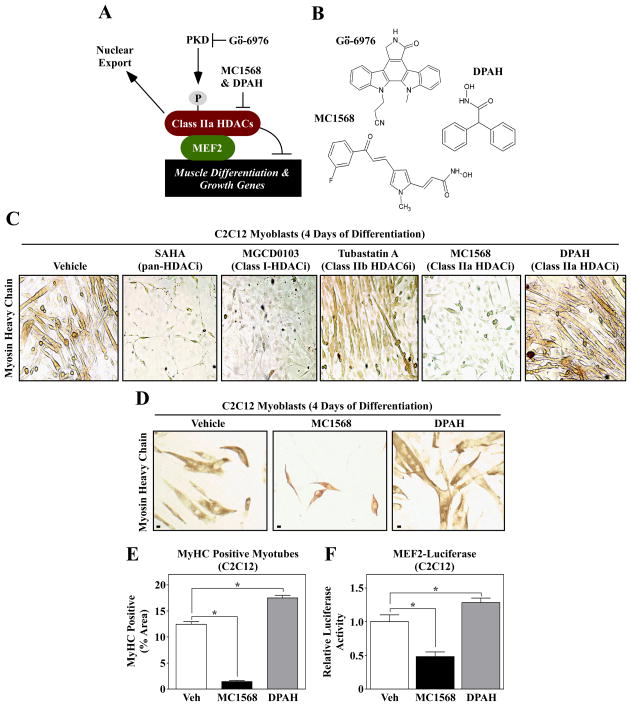
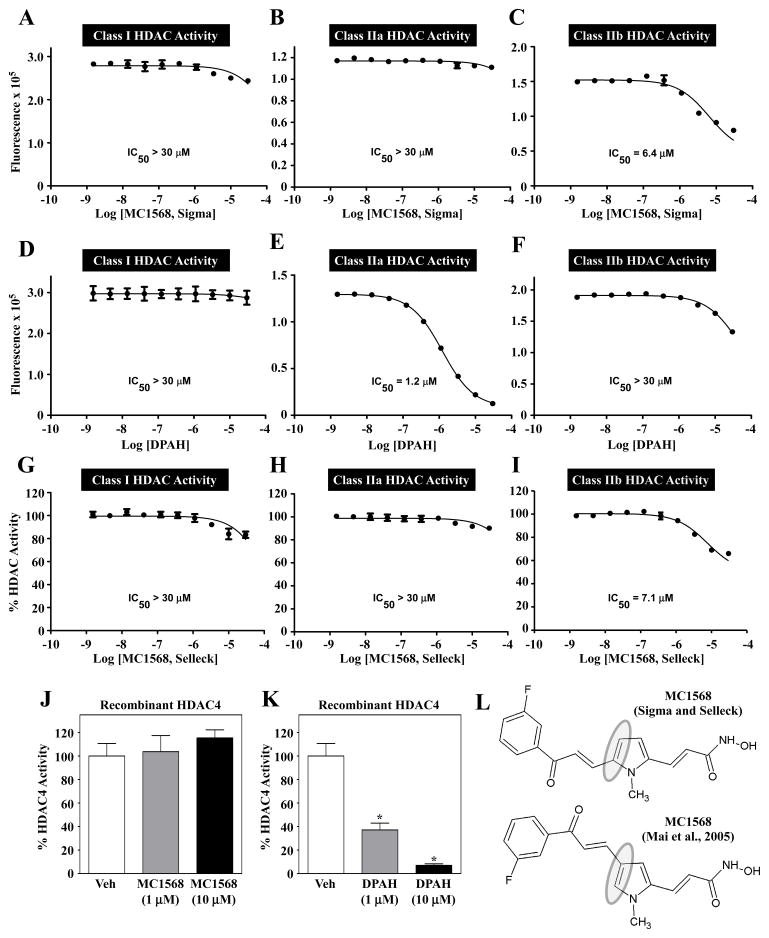
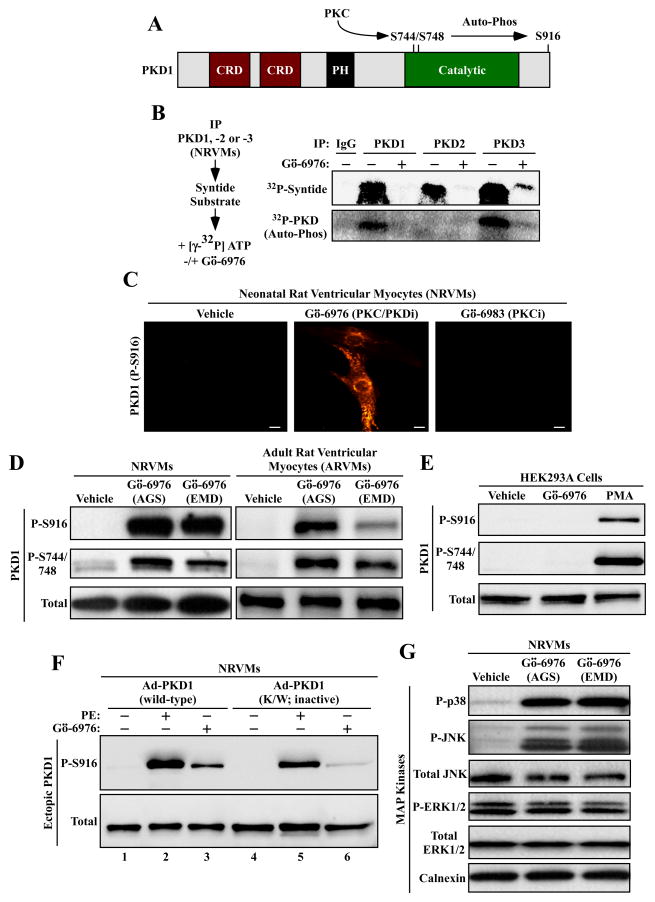
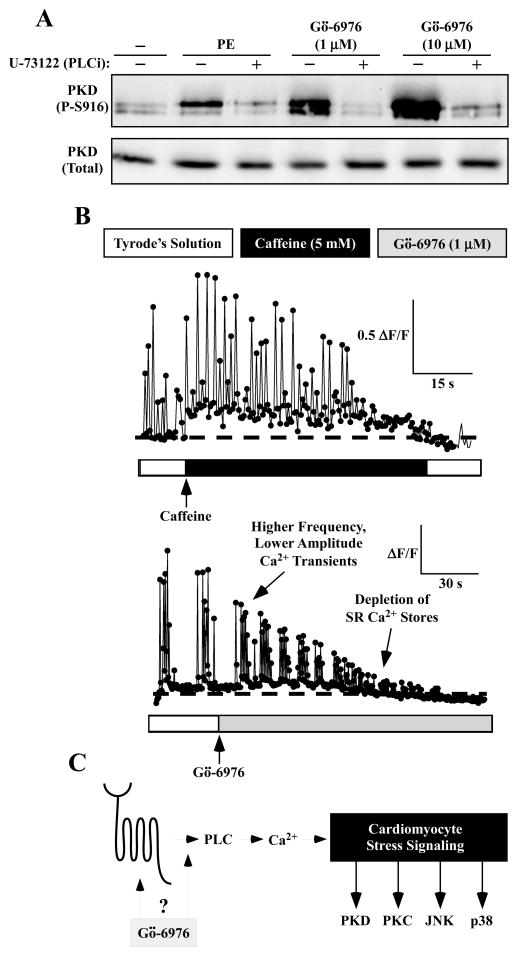
PKD-mediated phosphorylation of class IIa HDACs frees the MEF2 transcription factor to activate genes that govern muscle differentiation and growth. Studies of the regulation and function of this signaling axis have involved MC1568 and Gö-6976, which are small molecule inhibitors of class IIa HDAC and PKD catalytic activity, respectively. We describe unanticipated effects of these compounds. MC1568 failed to inhibit class IIa HDAC catalytic activity in vitro, and exerted divergent effects on skeletal muscle differentiation compared to a bona fide inhibitor of these HDACs. In cardiomyocytes, Gö-6976 triggered calcium signaling and activated stress-inducible kinases. Based on these findings, caution is warranted when employing MC1568 and Gö-6976 as pharmacological tool compounds to assess functions of class IIa HDACs and PKD.
Keywords: Class IIa histone deacetylase; Muscle; Protein kinase D; Small molecule inhibitors.
Copyright © 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Gregoretti IV, Lee YM, Goodson HV. Molecular evolution of the histone deacetylase family: functional implications of phylogenetic analysis. J Mol Biol. 2004;338:17–31. - PubMed
-
- McKinsey TA, Zhang CL, Olson EN. Signaling chromatin to make muscle. Curr Opin Cell Biol. 2002;14:763–772. - PubMed
-
- Lemercier C, Verdel A, Galloo B, Curtet S, Brocard MP, Khochbin S. mHDA1/HDAC5 histone deacetylase interacts with and represses MEF2A transcriptional activity. J Biol Chem. 2000;275:15594–15599. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
