

Structural basis for molecular discrimination by a 3',3'-cGAMP sensing riboswitch
- PMID: 25818298
- PMCID: PMC4732562
- DOI: 10.1016/j.celrep.2015.03.004
Structural basis for molecular discrimination by a 3',3'-cGAMP sensing riboswitch
Erratum in
- Cell Rep. 2015 Apr 28;11(4):671
Abstract
Cyclic dinucleotides are second messengers that target the adaptor STING and stimulate the innate immune response in mammals. Besides protein receptors, there are bacterial riboswitches that selectively recognize cyclic dinucleotides. We recently discovered a natural riboswitch that targets 3',3'-cGAMP, which is distinguished from the endogenous mammalian signal 2',3'-cGAMP by its backbone connectivity. Here, we report on structures of the aptamer domain of the 3',3'-cGAMP riboswitch from Geobacter in the 3',3'-cGAMP and c-di-GMP bound states. The riboswitch adopts a tuning fork-like architecture with a junctional ligand-binding pocket and different orientations of the arms are correlated with the identity of the bound cyclic dinucleotide. Subsequent biochemical experiments revealed that specificity of ligand recognition can be affected by point mutations outside of the binding pocket, which has implications for both the assignment and reengineering of riboswitches in this structural class.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
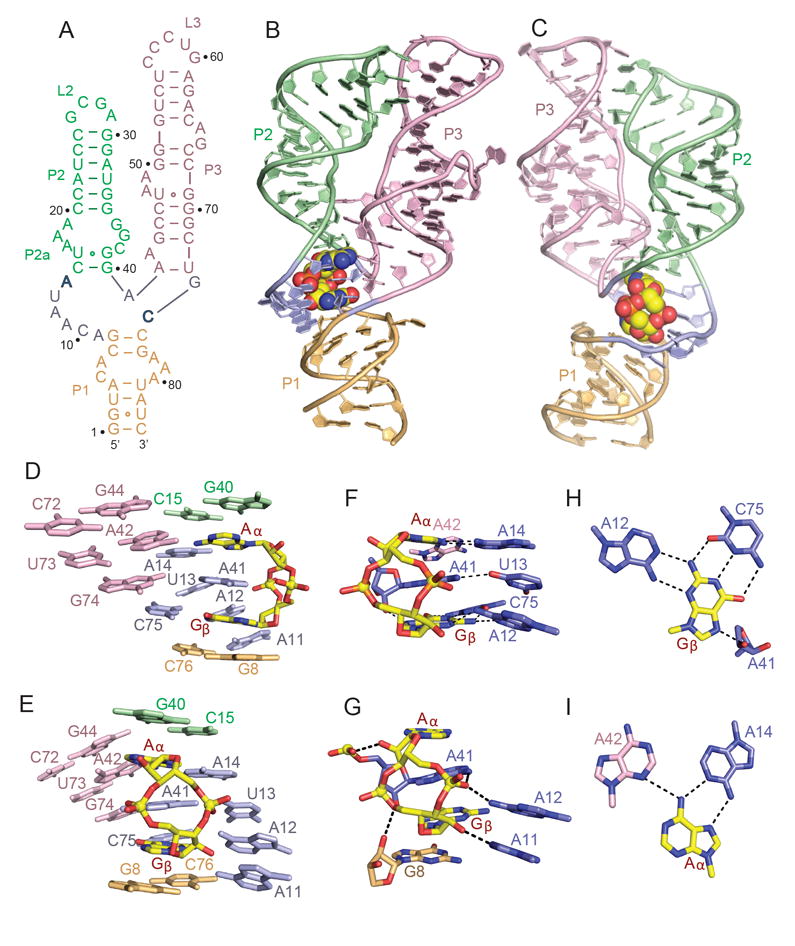
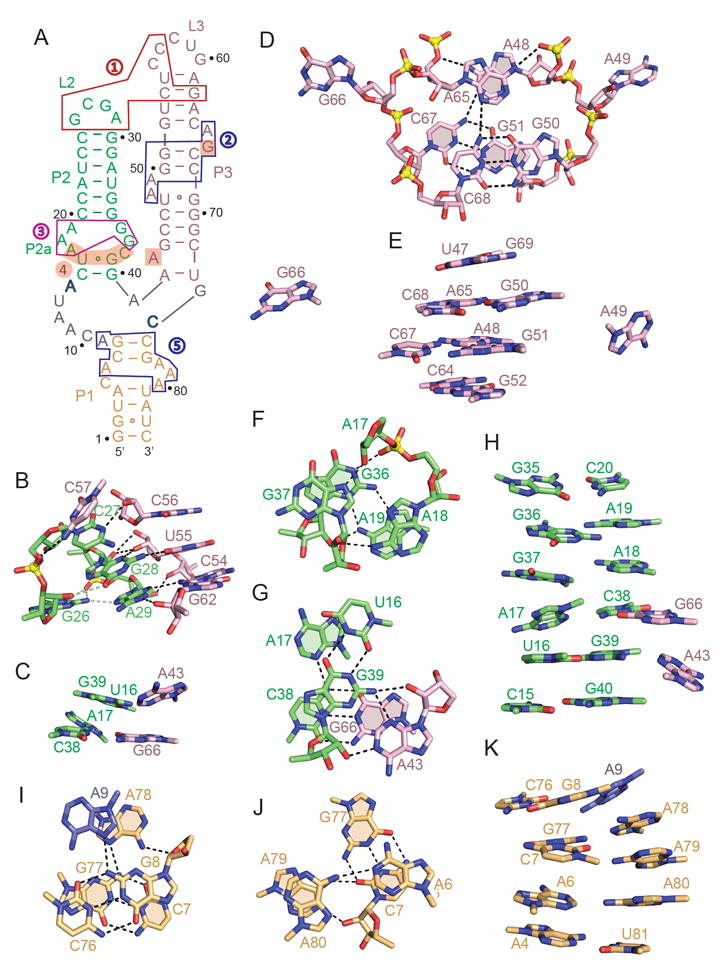
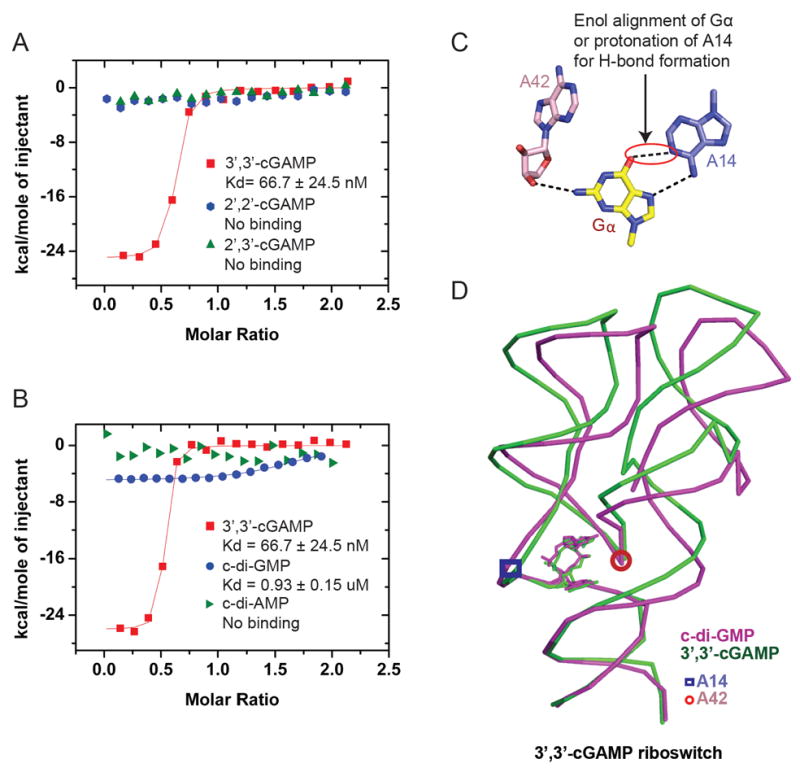
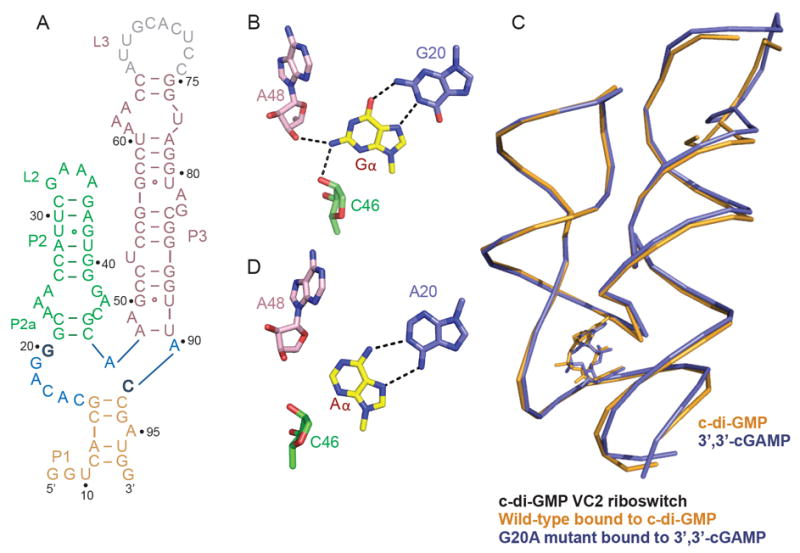
References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Loerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr. 2002:1948–1954. - PubMed
-
- Batey RT, Gilbert SD, Montange RK. Structure of a natural guanine-responsive riboswitch complexed with the metabolite hypoxanthine. Nature. 2004;432:411–415. - PubMed
-
- Cai X, Chiu YH, Chen ZJ. The cGAS-cGAMP-STING pathway of cytosolic DNA sensing and signaling. Mol Cell. 2014;54:289–296. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
