

The dynamic sclera: extracellular matrix remodeling in normal ocular growth and myopia development
- PMID: 25819458
- PMCID: PMC4379420
- DOI: 10.1016/j.exer.2014.07.015
The dynamic sclera: extracellular matrix remodeling in normal ocular growth and myopia development
Abstract
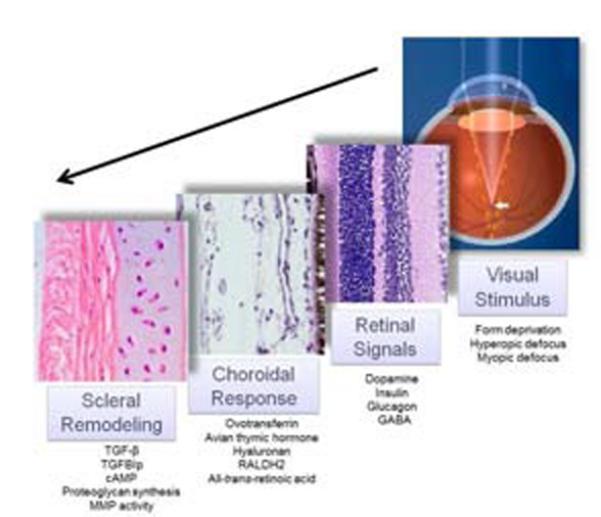
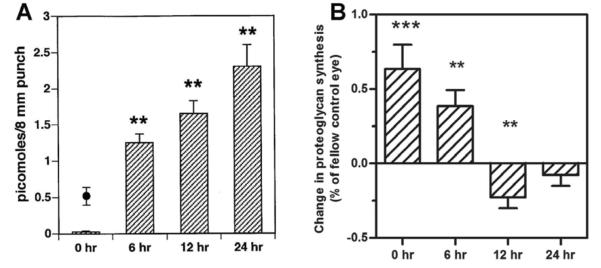
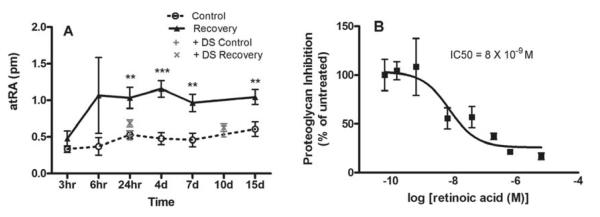
Myopia is a common ocular condition, characterized by excessive elongation of the ocular globe. The prevalence of myopia continues to increase, particularly among highly educated groups, now exceeding 80% in some groups. In parallel with the increased prevalence of myopia, are increases in associated blinding ocular conditions including glaucoma, retinal detachment and macular degeneration, making myopia a significant global health concern. The elongation of the eye is closely related to the biomechanical properties of the sclera, which in turn are largely dependent on the composition of the scleral extracellular matrix. Therefore an understanding of the cellular and extracellular events involved in the regulation of scleral growth and remodeling during childhood and young adulthood will provide future avenues for the treatment of myopia and its associated ocular complications.
Keywords: emmetropization; extracellular matrix; myopia; ocular development; sclera.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures


References
-
- Annunen S, Korkko J, Czarny M, Warman ML, Brunner HG, Kaariainen H, Mulliken JB, Tranebjaerg L, Brooks DG, Cox GF, Cruysberg JR, Curtis MA, Davenport SL, Friedrich CA, Kaitila I, Krawczynski MR, Latos-Bielenska A, Mukai S, Olsen BR, Shinno N, Somer M, Vikkula M, Zlotogora J, Prockop DJ, Ala-Kokko L. Splicing mutations of 54-bp exons in the COL11A1 gene cause Marshall syndrome, but other mutations cause overlapping Marshall/Stickler phenotypes. Am J Hum Genet. 1999;65:974–983. - PMC - PubMed
-
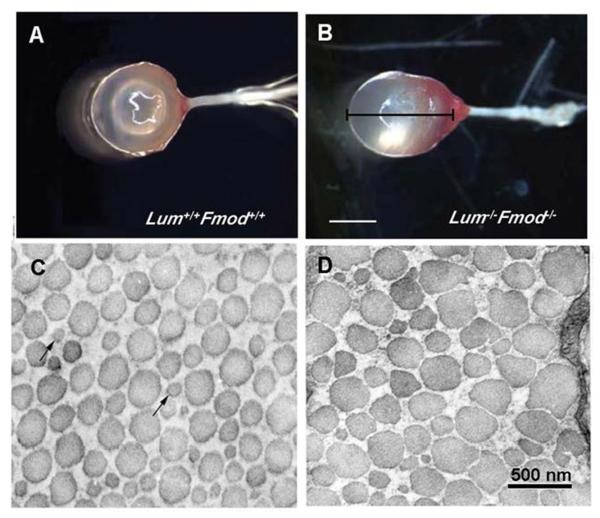
- Austin BA, Coulon C, Liu CY, Kao WW, Rada JA. Altered collagen fibril formation in the sclera of lumican-deficient mice. Invest Ophthalmol Vis Sci. 2002;43:1695–1701. - PubMed
-
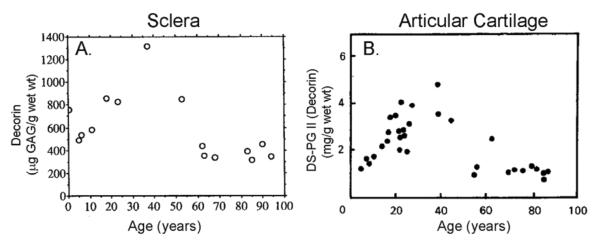
- Avetisov ES, Savitskaya NF, Vinetskaya MI, Iomdina EN. A study of biochemical and biomechanical qualities of normal and myopic eye sclera in humans of different age groups. Metab Pediatr Syst Ophthalmol. 1983;7:183–188. - PubMed
-
- Barathi VA, Boopathi VG, Yap EP, Beuerman RW. Two models of experimental myopia in the mouse. Vision Res. 2008;48:904–916. - PubMed
-
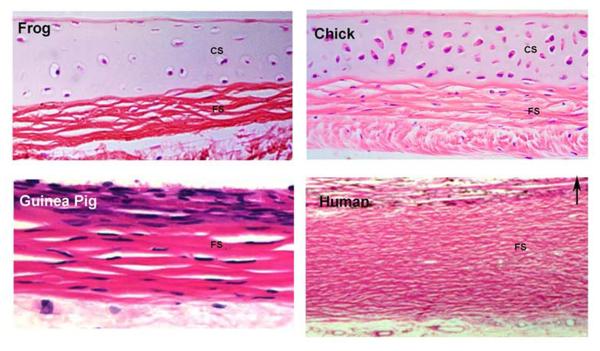
- Birk DE, Lande MA. Corneal and scleral collagen fiber formation in vitro. Biochim Biophys Acta. 1981;670:362–369. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
