

Modular deconstruction reveals the dynamical and physical building blocks of a locomotion motor program
- PMID: 25819612
- PMCID: PMC6016739
- DOI: 10.1016/j.neuron.2015.03.005
Modular deconstruction reveals the dynamical and physical building blocks of a locomotion motor program
Abstract
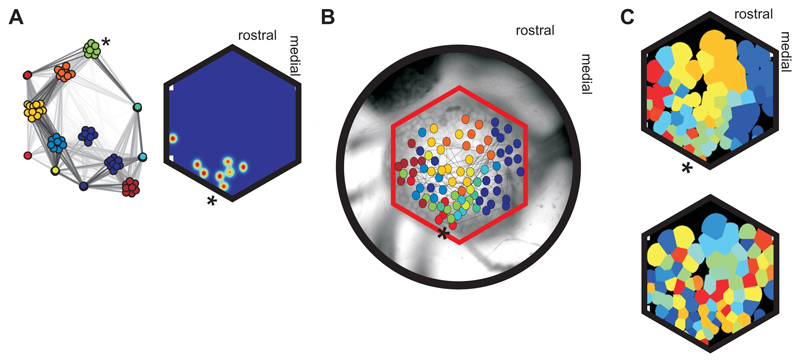
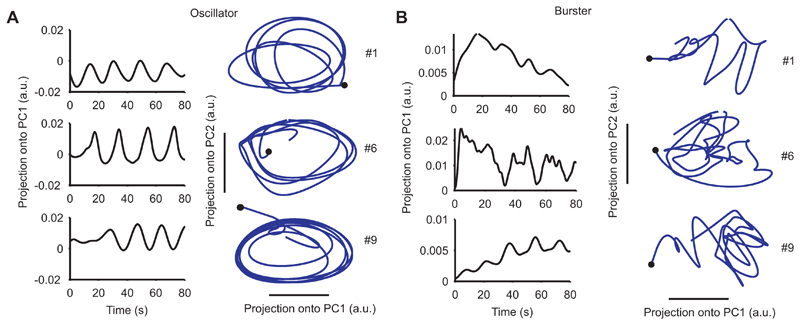
The neural substrates of motor programs are only well understood for small, dedicated circuits. Here we investigate how a motor program is constructed within a large network. We imaged populations of neurons in the Aplysia pedal ganglion during execution of a locomotion motor program. We found that the program was built from a very small number of dynamical building blocks, including both neural ensembles and low-dimensional rotational dynamics. These map onto physically discrete regions of the ganglion, so that the motor program has a corresponding modular organization in both dynamical and physical space. Using this dynamic map, we identify the population potentially implementing the rhythmic pattern generator and find that its activity physically traces a looped trajectory, recapitulating its low-dimensional rotational dynamics. Our results suggest that, even in simple invertebrates, neural motor programs are implemented by large, distributed networks containing multiple dynamical systems.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Unraveling a locomotor network, many neurons at a time.Neuron. 2015 Apr 8;86(1):9-11. doi: 10.1016/j.neuron.2015.03.056. Neuron. 2015. PMID: 25856478
References
-
- Ahrens MB, Orger MB, Robson DN, Li JM, Keller PJ. Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat Methods. 2013;10:413–420. - PubMed
-
- Berg RW, Alaburda A, Hounsgaard J. Balanced inhibition and excitation drive spike activity in spinal half-centers. Science. 2007;315:390–393. - PubMed
-
- Brezina V, Orekhova IV, Weiss KR. The neuromuscular transform: the dynamic, nonlinear link between motor neuron firing patterns and muscle contraction in rhythmic behaviors. J Neurophysiol. 2000;83:207–231. - PubMed
-
- Briggman KL, Abarbanel HDI, Kristan W., Jr Optical imaging of neuronal populations during decision-making. Science. 2005;307:896–901. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
