

Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-β
- PMID: 25822997
- PMCID: PMC4379073
- DOI: 10.1371/journal.pone.0121123
Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-β
Abstract
Scope: Extracellular vesicles, including exosomes, have been identified in all biological fluids and rediscovered as an important part of the intercellular communication. Breast milk also contains extracellular vesicles and the proposed biological function is to enhance the antimicrobial defense in newborns. It is, however, unknown whether extracellular vesicles are still present in commercial milk and, more importantly, whether they retained their bioactivity. Here, we characterize the extracellular vesicles present in semi-skimmed cow milk available for consumers and study their effect on T cells.
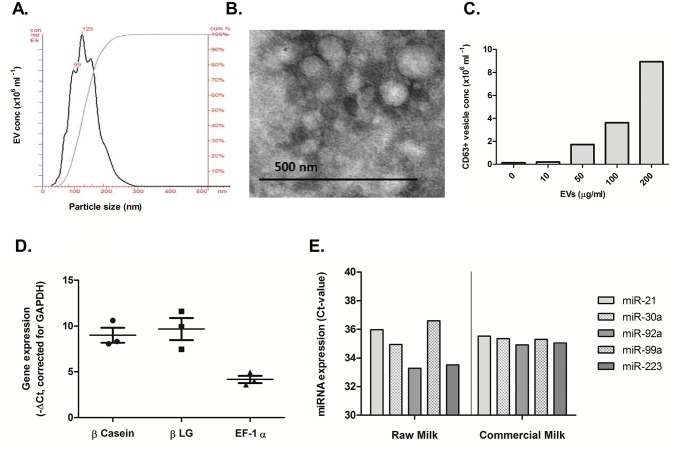
Methods and results: Extracellular vesicles from commercial milk were isolated and characterized. Milk-derived extracellular vesicles contained several immunomodulating miRNAs and membrane protein CD63, characteristics of exosomes. In contrast to RAW 267.4 derived extracellular vesicles the milk-derived extracellular vesicles were extremely stable under degrading conditions, including low pH, boiling and freezing. Milk-derived extracellular vesicles were easily taken up by murine macrophages in vitro. Furthermore, we found that they can facilitate T cell differentiation towards the pathogenic Th17 lineage. Using a (CAGA)12-luc reporter assay we showed that these extracellular vesicles carried bioactive TGF-β, and that anti-TGF-β antibodies blocked Th17 differentiation.
Conclusion: Our findings show that commercial milk contains stable extracellular vesicles, including exosomes, and carry immunoregulatory cargo. These data suggest that the extracellular vesicles present in commercial cow milk remains intact in the gastrointestinal tract and exert an immunoregulatory effect.
Conflict of interest statement
Figures



Similar articles
-
Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models.Mol Nutr Food Res. 2015 Sep;59(9):1701-12. doi: 10.1002/mnfr.201500222. Epub 2015 Jul 1. Mol Nutr Food Res. 2015. PMID: 26047123
-
Isolation and Characterization of Cow-, Buffalo-, Sheep- and Goat-Milk-Derived Extracellular Vesicles.Cells. 2023 Oct 20;12(20):2491. doi: 10.3390/cells12202491. Cells. 2023. PMID: 37887335 Free PMC article.
-
Yak-milk-derived exosomes promote proliferation of intestinal epithelial cells in an hypoxic environment.J Dairy Sci. 2019 Feb;102(2):985-996. doi: 10.3168/jds.2018-14946. Epub 2018 Dec 20. J Dairy Sci. 2019. PMID: 30580945
-
Flood Control: How Milk-Derived Extracellular Vesicles Can Help to Improve the Intestinal Barrier Function and Break the Gut-Joint Axis in Rheumatoid Arthritis.Front Immunol. 2021 Jul 28;12:703277. doi: 10.3389/fimmu.2021.703277. eCollection 2021. Front Immunol. 2021. PMID: 34394100 Free PMC article. Review.
-
Regulation of T Cell Activation and Differentiation by Extracellular Vesicles and Their Pathogenic Role in Systemic Lupus Erythematosus and Multiple Sclerosis.Molecules. 2017 Feb 2;22(2):225. doi: 10.3390/molecules22020225. Molecules. 2017. PMID: 28157168 Free PMC article. Review.
Cited by
-
MiR-21: an environmental driver of malignant melanoma?J Transl Med. 2015 Jun 27;13:202. doi: 10.1186/s12967-015-0570-5. J Transl Med. 2015. PMID: 26116372 Free PMC article. Review.
-
Ameliorative Effects of Camel Milk and Its Exosomes on Diabetic Nephropathy in Rats.Membranes (Basel). 2022 Oct 28;12(11):1060. doi: 10.3390/membranes12111060. Membranes (Basel). 2022. PMID: 36363614 Free PMC article.
-
Small RNAs in eucaryotes: new clues for amplifying microRNA benefits.Cell Biosci. 2020 Jan 3;10:1. doi: 10.1186/s13578-019-0370-3. eCollection 2020. Cell Biosci. 2020. PMID: 31911829 Free PMC article. Review.
-
Bovine Milk Extracellular Vesicles (EVs) Modification Elicits Skeletal Muscle Growth in Rats.Front Physiol. 2019 Apr 16;10:436. doi: 10.3389/fphys.2019.00436. eCollection 2019. Front Physiol. 2019. PMID: 31040795 Free PMC article.
-
Latest Trend of Milk Derived Exosomes: Cargos, Functions, and Applications.Front Nutr. 2021 Oct 29;8:747294. doi: 10.3389/fnut.2021.747294. eCollection 2021. Front Nutr. 2021. PMID: 34778341 Free PMC article. Review.
References
-
- Théry C, Zitvogel L, Amigorena S (2002) Exosomes: composition, biogenesis and function. Nature Reviews Immunology 2: 569–579. - PubMed
-
- Blanchard N, Lankar D, Faure F, Regnault A, Dumont C, Raposo G, et al. (2002) TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/ζ complex. The Journal of Immunology 168: 3235–3241. - PubMed
-
- Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, et al. (1998) Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell derived exosomes. Nature medicine 4: 594–600. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
