

T cell lipid peroxidation induces ferroptosis and prevents immunity to infection
- PMID: 25824823
- PMCID: PMC4387287
- DOI: 10.1084/jem.20140857
T cell lipid peroxidation induces ferroptosis and prevents immunity to infection
Abstract
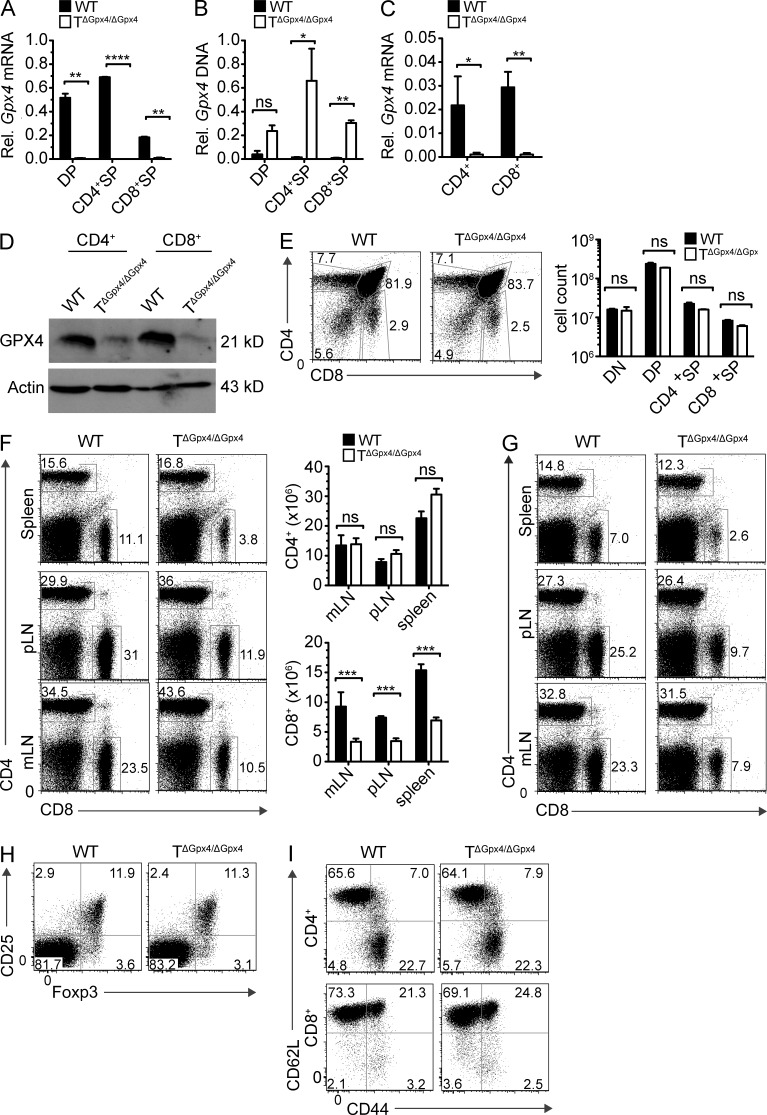
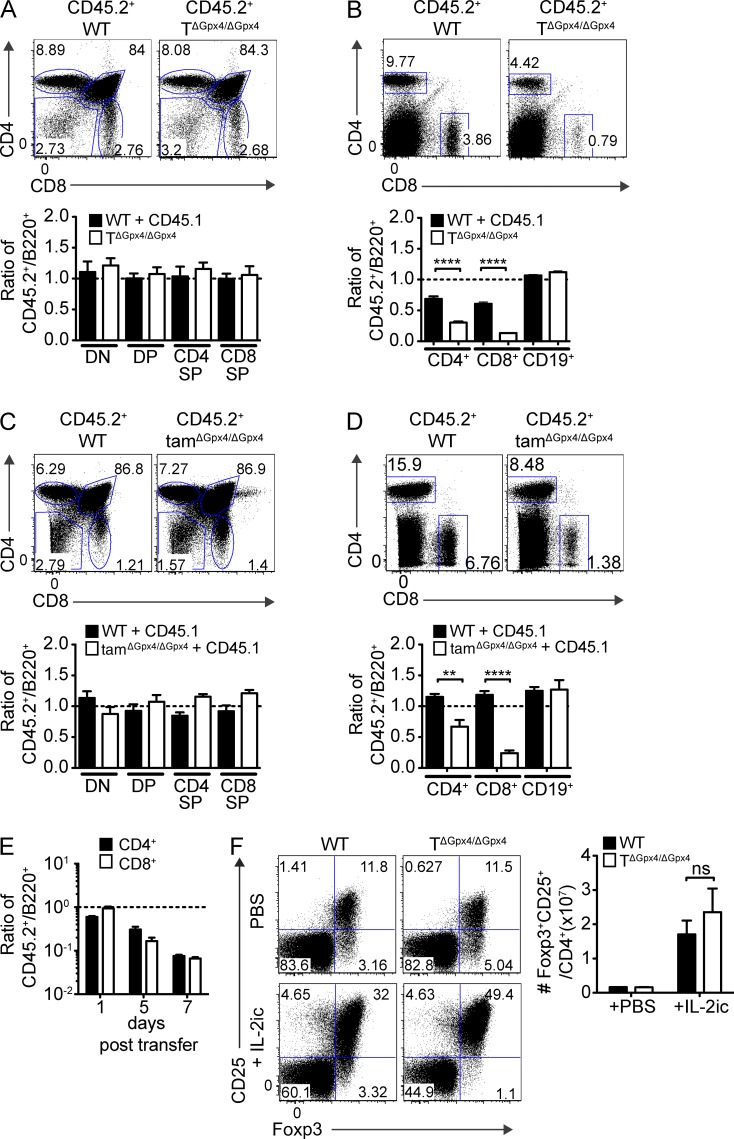
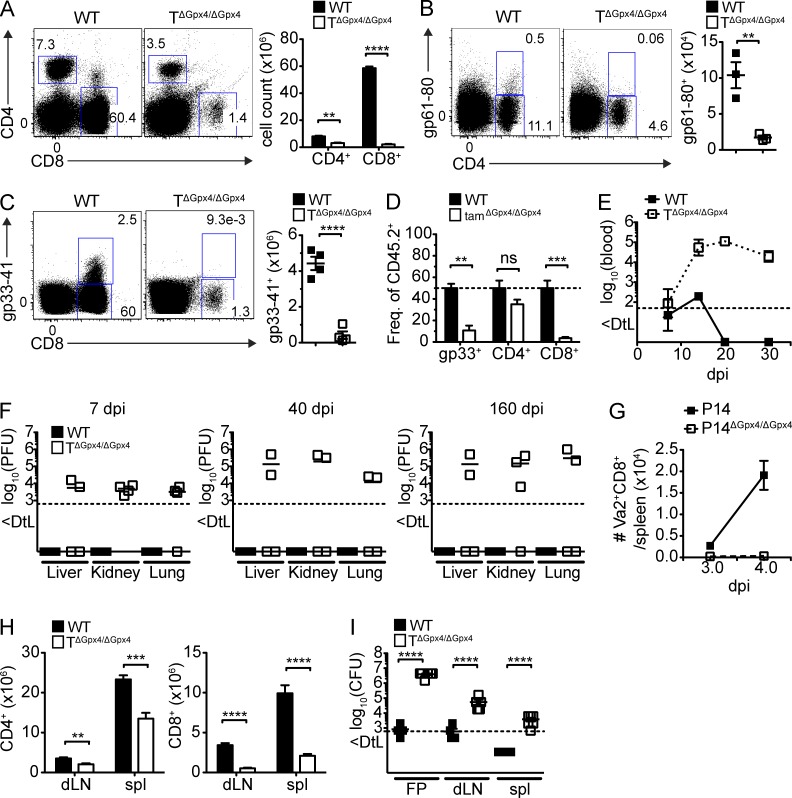
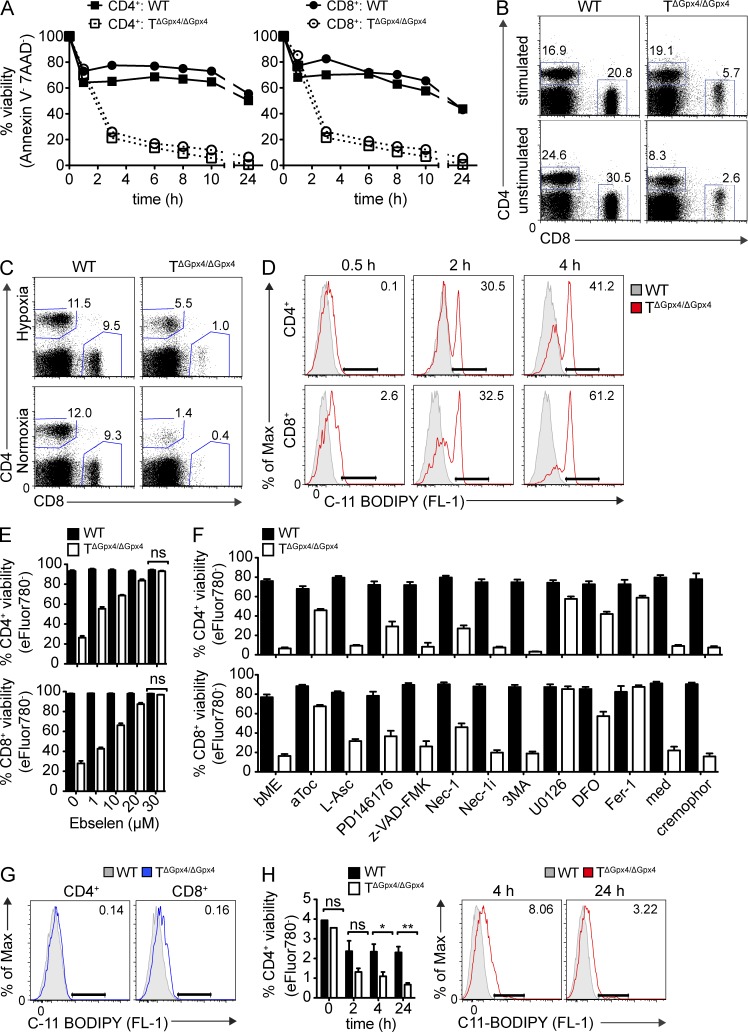
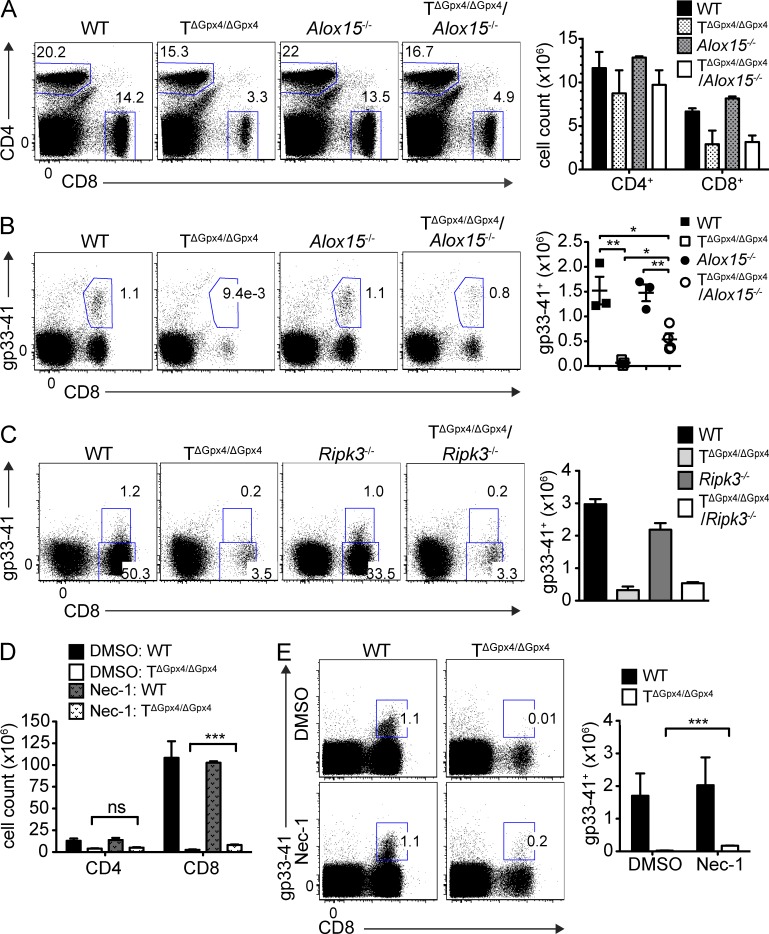
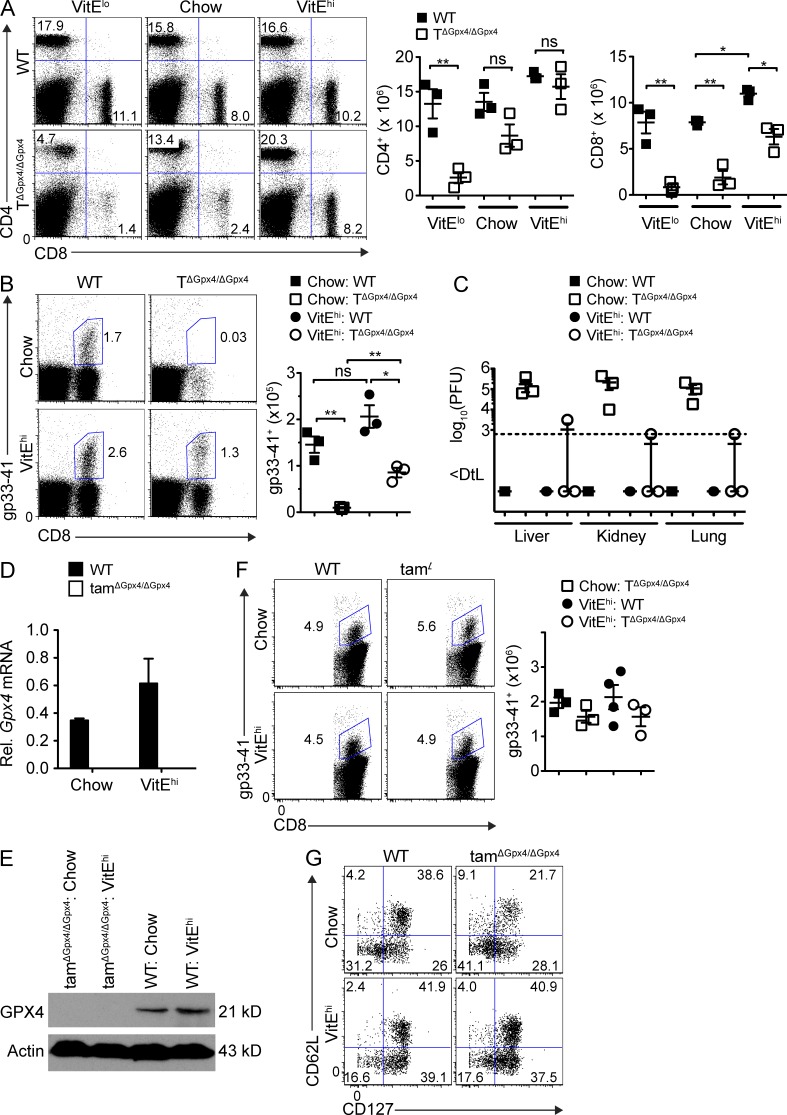
The selenoenzyme glutathione peroxidase 4 (Gpx4) is a major scavenger of phospholipid hydroperoxides. Although Gpx4 represents a key component of the reactive oxygen species-scavenging network, its relevance in the immune system is yet to be defined. Here, we investigated the importance of Gpx4 for physiological T cell responses by using T cell-specific Gpx4-deficient mice. Our results revealed that, despite normal thymic T cell development, CD8(+) T cells from T(ΔGpx4/ΔGpx4) mice had an intrinsic defect in maintaining homeostatic balance in the periphery. Moreover, both antigen-specific CD8(+) and CD4(+) T cells lacking Gpx4 failed to expand and to protect from acute lymphocytic choriomeningitis virus and Leishmania major parasite infections, which were rescued with diet supplementation of high dosage of vitamin E. Notably, depletion of the Gpx4 gene in the memory phase of viral infection did not affect T cell recall responses upon secondary infection. Ex vivo, Gpx4-deficient T cells rapidly accumulated membrane lipid peroxides and concomitantly underwent cell death driven by ferroptosis but not necroptosis. These studies unveil an essential role of Gpx4 for T cell immunity.
© 2015 Matsushita et al.
Figures
References
-
- Angelini G., Gardella S., Ardy M., Ciriolo M.R., Filomeni G., Di Trapani G., Clarke F., Sitia R., and Rubartelli A.. 2002. Antigen-presenting dendritic cells provide the reducing extracellular microenvironment required for T lymphocyte activation. Proc. Natl. Acad. Sci. USA. 99:1491–1496 10.1073/pnas.022630299 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
