

Advances in Understanding Carboxysome Assembly in Prochlorococcus and Synechococcus Implicate CsoS2 as a Critical Component
- PMID: 25826651
- PMCID: PMC4499774
- DOI: 10.3390/life5021141
Advances in Understanding Carboxysome Assembly in Prochlorococcus and Synechococcus Implicate CsoS2 as a Critical Component
Abstract
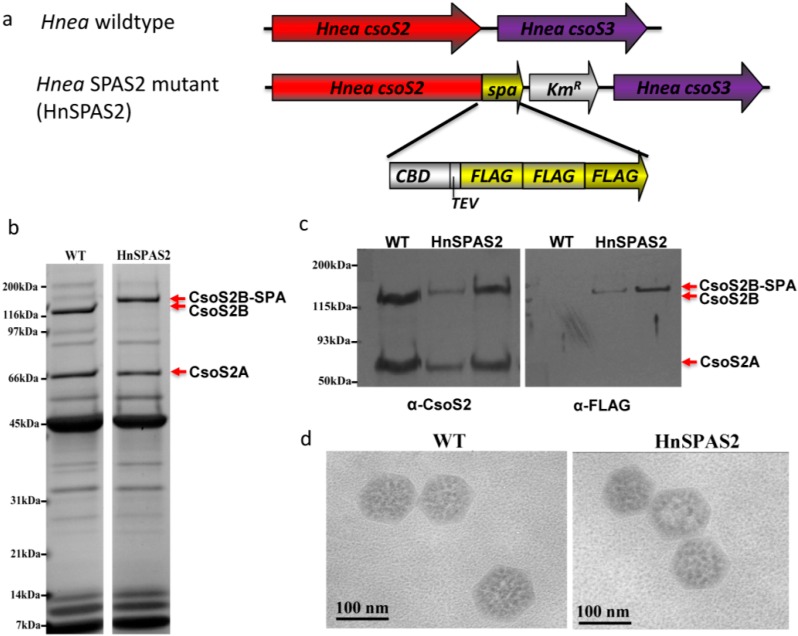
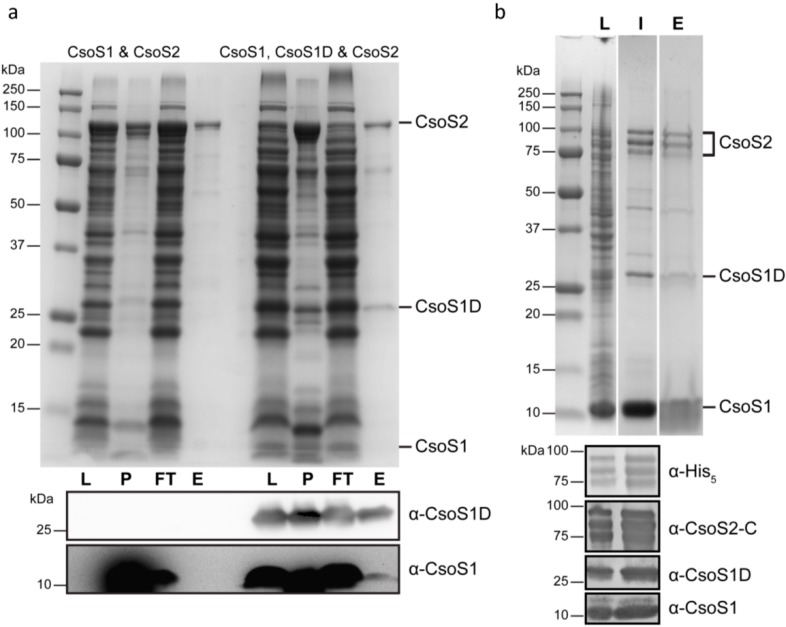
The marine Synechococcus and Prochlorococcus are the numerically dominant cyanobacteria in the ocean and important in global carbon fixation. They have evolved a CO2-concentrating-mechanism, of which the central component is the carboxysome, a self-assembling proteinaceous organelle. Two types of carboxysome, α and β, encapsulating form IA and form IB d-ribulose-1,5-bisphosphate carboxylase/oxygenase, respectively, differ in gene organization and associated proteins. In contrast to the β-carboxysome, the assembly process of the α-carboxysome is enigmatic. Moreover, an absolutely conserved α-carboxysome protein, CsoS2, is of unknown function and has proven recalcitrant to crystallization. Here, we present studies on the CsoS2 protein in three model organisms and show that CsoS2 is vital for α-carboxysome biogenesis. The primary structure of CsoS2 appears tripartite, composed of an N-terminal, middle (M)-, and C-terminal region. Repetitive motifs can be identified in the N- and M-regions. Multiple lines of evidence suggest CsoS2 is highly flexible, possibly an intrinsically disordered protein. Based on our results from bioinformatic, biophysical, genetic and biochemical approaches, including peptide array scanning for protein-protein interactions, we propose a model for CsoS2 function and its spatial location in the α-carboxysome. Analogies between the pathway for β-carboxysome biogenesis and our model for α-carboxysome assembly are discussed.
Figures










References
-
- Goericke R., Welschmeyer N.A. The marine prochlorophyte Prochlorococcus contributes significantly to phytoplankton biomass and primary production in the Sargasso Sea. Deep Sea Res. 1993;40:2283–2294. doi: 10.1016/0967-0637(93)90104-B. - DOI
-
- Liu H., Nolla H.A., Campbell L. Prochlorococcus growth rate and contribution to primary production in the equatorial and subtropical North Pacific Ocean. Aquat. Microb. Ecol. 1997;12:39–47. doi: 10.3354/ame012039. - DOI
-
- Liu H., Campbell L., Landry M.R., Nolla H.A., Brown S.L., Constantinou J. Prochlorococcus and Synechococcus growth rates and contributions to production in the Arabian Sea during the 1995 Southwest and Northeast Monsoons. Deep Sea Res. 1998;45:2327–2352. doi: 10.1016/S0967-0645(98)00073-3. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
