

miR-214 promotes osteoclastogenesis by targeting Pten/PI3k/Akt pathway
- PMID: 25826666
- PMCID: PMC4615895
- DOI: 10.1080/15476286.2015.1017205
miR-214 promotes osteoclastogenesis by targeting Pten/PI3k/Akt pathway
Abstract
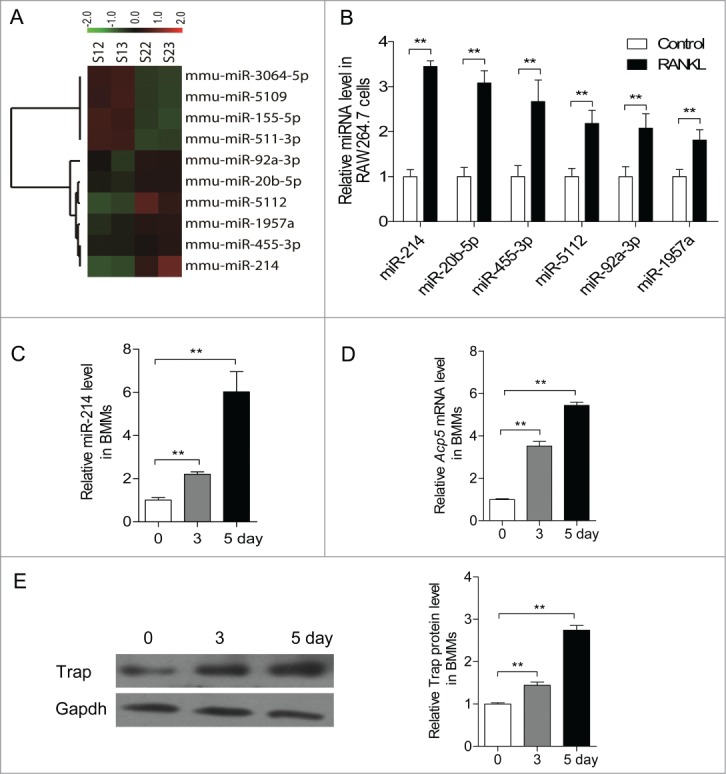
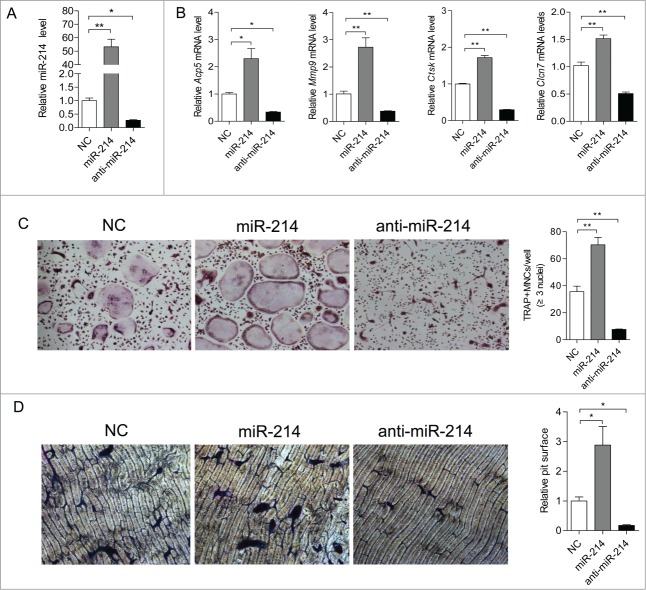
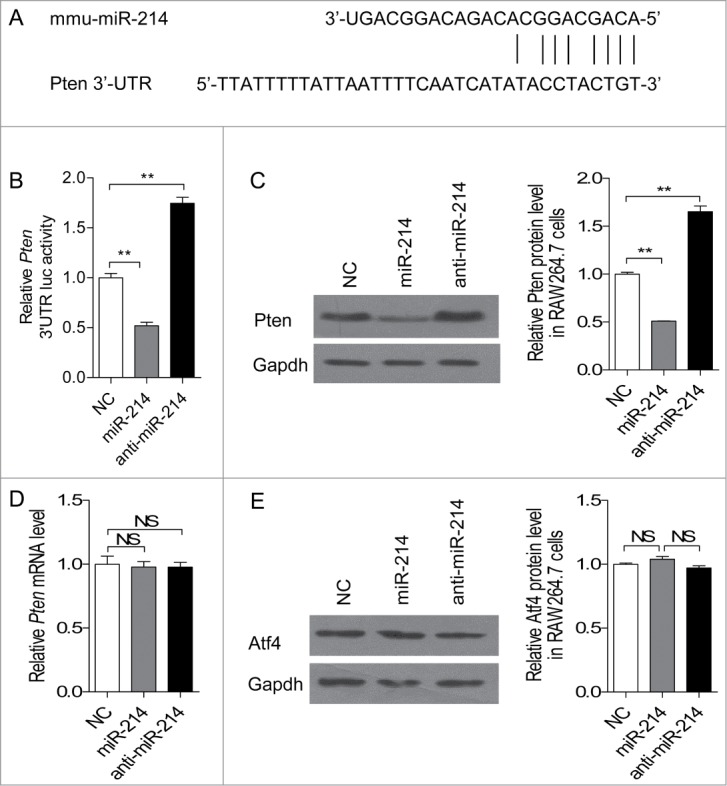
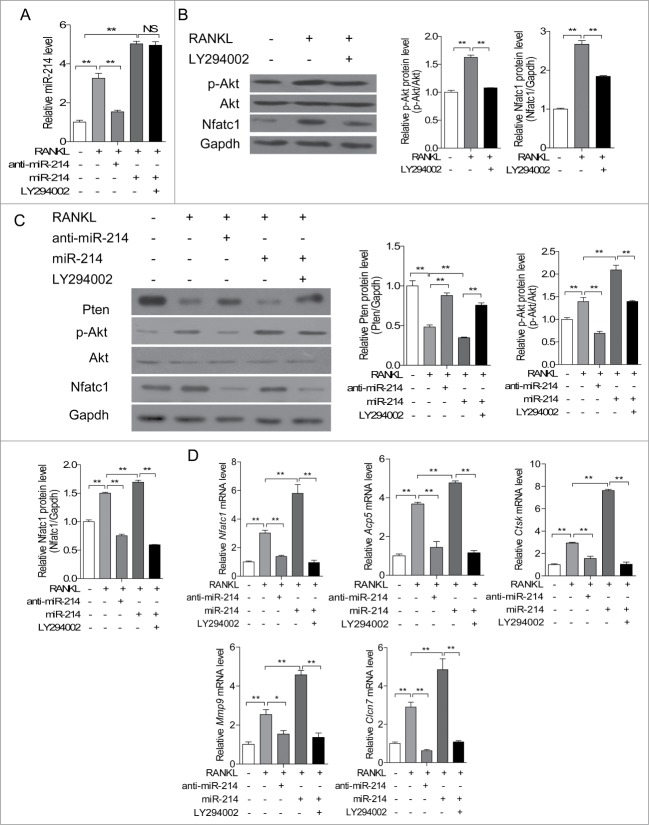
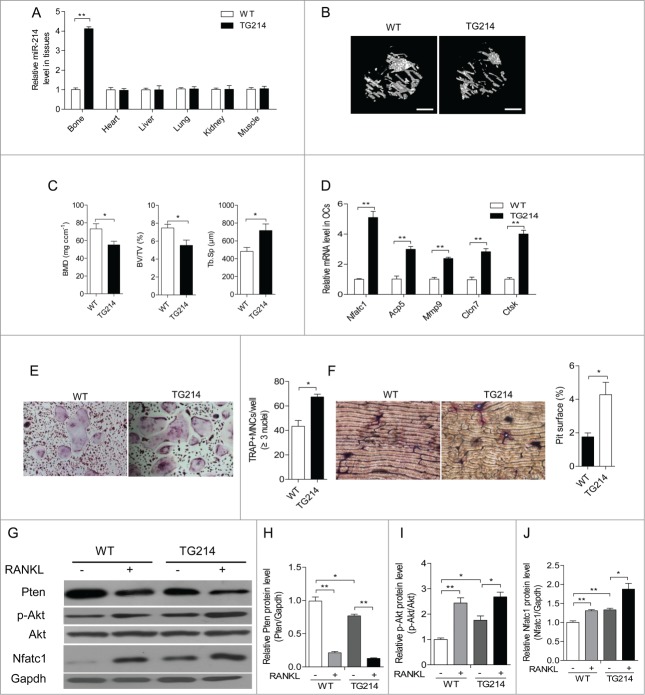
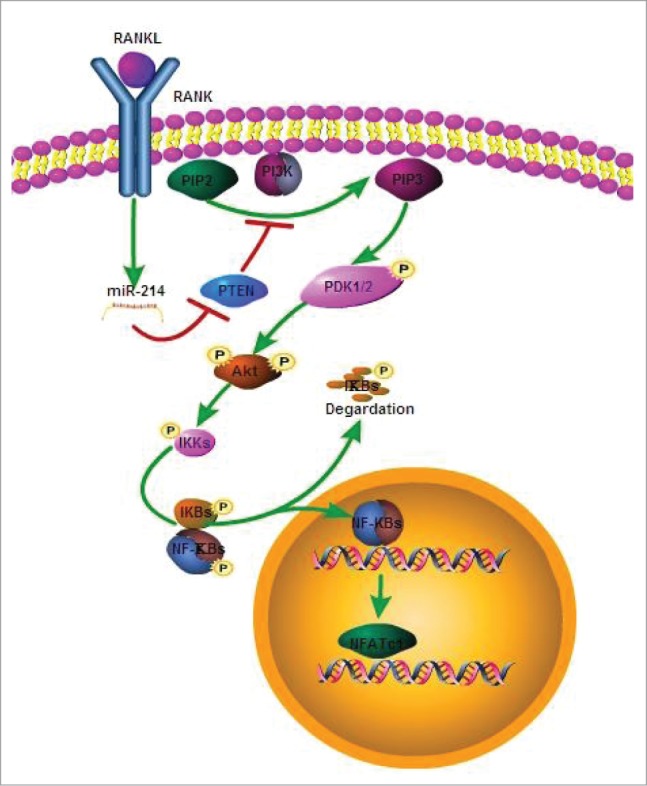
microRNA is necessary for osteoclast differentiation, function and survival. It has been reported that miR-199/214 cluster plays important roles in vertebrate skeletal development and miR-214 inhibits osteoblast function by targeting ATF4. Here, we show that miR-214 is up-regulated during osteoclastogenesis from bone marrow monocytes (BMMs) with macrophage colony stimulating factor (M-CSF) and receptor activator of nuclear factor-κB ligand (RANKL) induction, which indicates that miR-214 plays a critical role in osteoclast differentiation. Overexpression of miR-214 in BMMs promotes osteoclastogenesis, whereas inhibition of miR-214 attenuates it. We further find that miR-214 functions through PI3K/Akt pathway by targeting phosphatase and tensin homolog (Pten). In vivo, osteoclast specific miR-214 transgenic mice (OC-TG214) exhibit down-regulated Pten levels, increased osteoclast activity, and reduced bone mineral density. These results reveal a crucial role of miR-214 in the differentiation of osteoclasts, which will provide a potential therapeutic target for osteoporosis.
Keywords: BMD, bone mineral density; BMMs, bone marrow monocytes; BV/TV, ratio of bone volume to tissue volume; Dnm3os, Dnm3 opposite strand; M-CSF, macrophage colony stimulating factor; NFATc1, nuclear factor of activated T-cells cytoplasmic; OC-TG214, osteoclast specific miR-214 transgenic mice; PI 3-kinase; PTEN; Pten, phosphatase and tensin homolog; RANKL, receptor activator of nuclear factor-κB ligand; TRAP, tartrate-resistant acid phosphatase; Tb.Sp, trabecular spacing; WT, wild-type; miRNA; micro CT, Micro computed tomography; osteoclast; osteoporosis; qRT-PCR, quantitative real-time PCR.
Figures
References
-
- Del Fattore A, Teti A, Rucci N. Osteoclast receptors and signaling. Arch Biochem Biophys 2008; 473:147–60; http://dx.doi.org/ 10.1016/j.abb.2008.01.011. - DOI - PubMed
-
- Ross FP, Teitelbaum SL. alphavbeta3 and macrophage colony-stimulating factor: partners in osteoclast biology. Immunol Rev 2005; 208:88–105; PMID:16313343; http://dx.doi.org/ 10.1111/j.0105-2896.2005.00331.x. - DOI - PubMed
-
- Miyazaki T, Tanaka S, Sanjay A, Baron R. The role of c-Src kinase in the regulation of osteoclast function. Mod Rheumatol 2006; 16: 68–74; PMID:16633924; http://dx.doi.org/ 10.3109/s10165-006-0460-z. - DOI - PubMed
-
- Edwards CM, Mundy GR. Eph receptors and ephrin signaling pathways: a role in bone homeostasis. Int J Med Sci 2008; 5:263–72; PMID:18797510; http://dx.doi.org/ 10.7150/ijms.5.263. - DOI - PMC - PubMed
-
- Hattersley G, Owens J, Flanagan AM, Chambers TJ. Macrophage colony stimulating factor (M-CSF) is essential for osteoclast formation in vitro. Biochem Biophys Res Commun 1991; 177:526–31; PMID:2043138; http://dx.doi.org/ 10.1016/0006-291X(91)92015-C. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous