

Genome sequence of the Drosophila melanogaster male-killing Spiroplasma strain MSRO endosymbiont
- PMID: 25827421
- PMCID: PMC4453565
- DOI: 10.1128/mBio.02437-14
Genome sequence of the Drosophila melanogaster male-killing Spiroplasma strain MSRO endosymbiont
Abstract
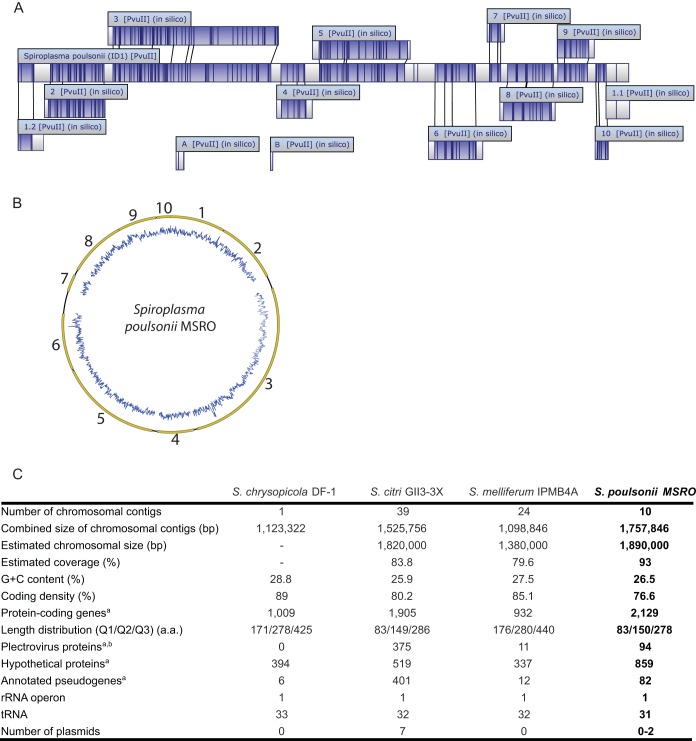
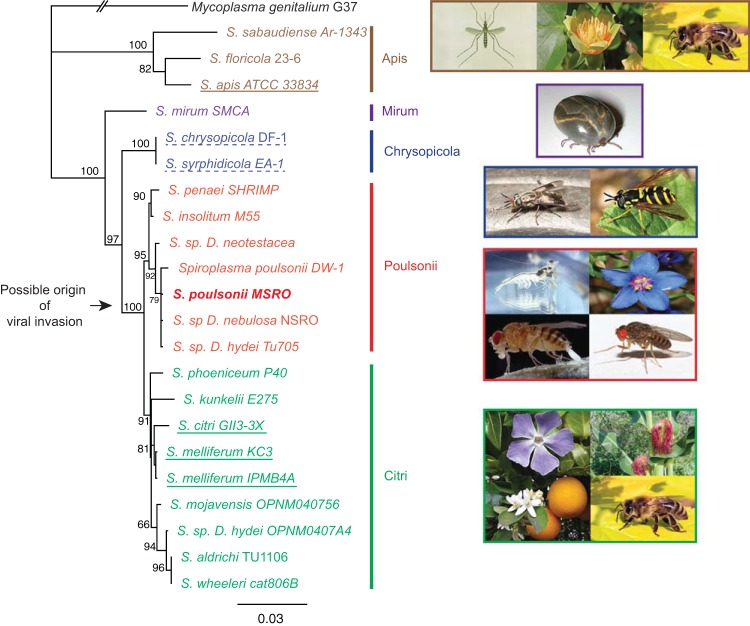
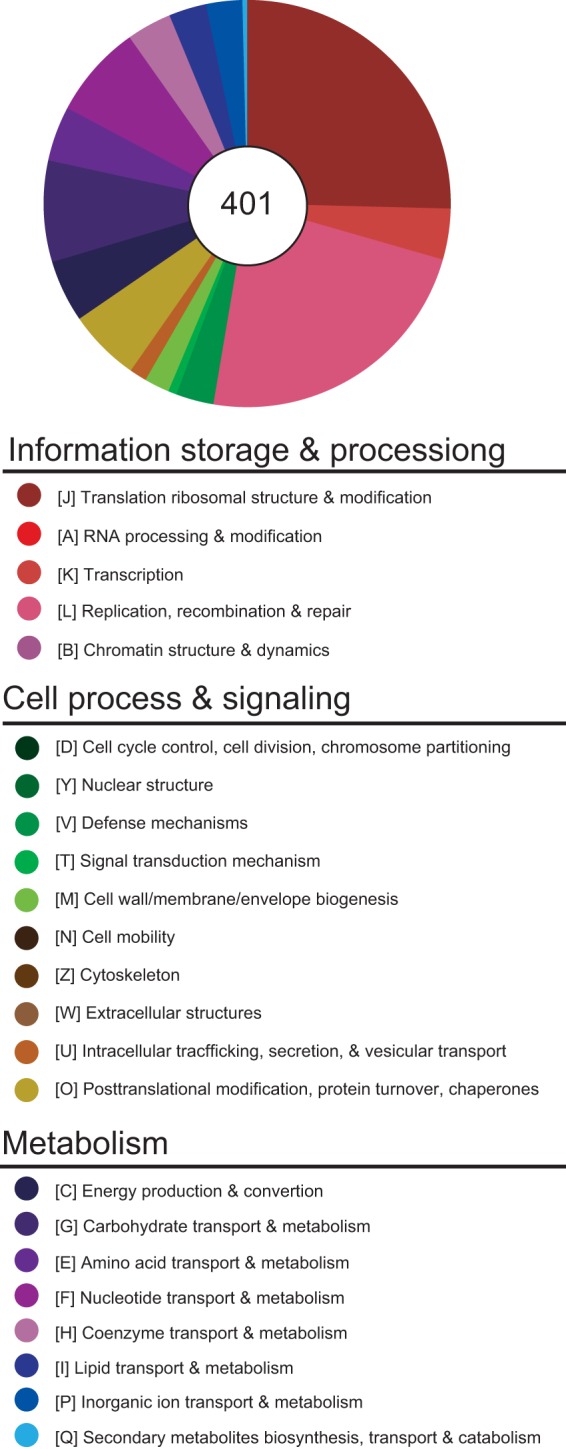
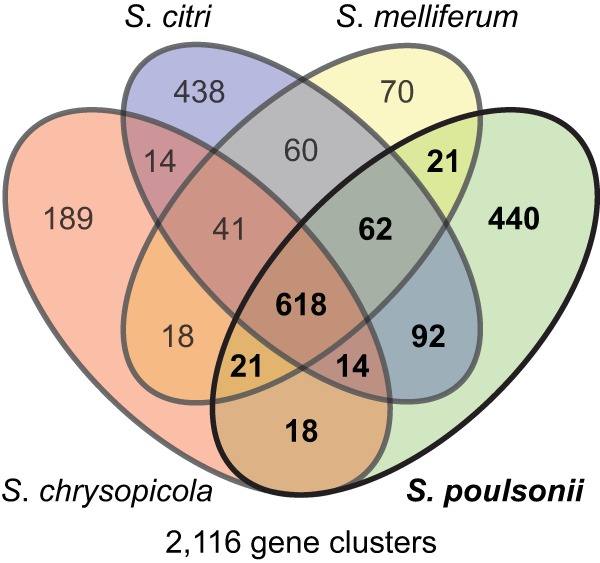
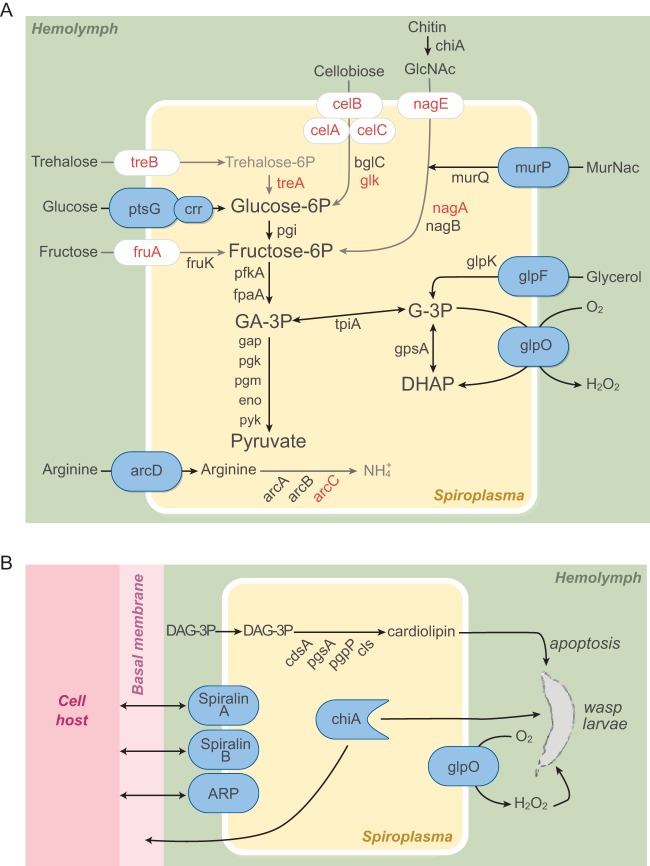
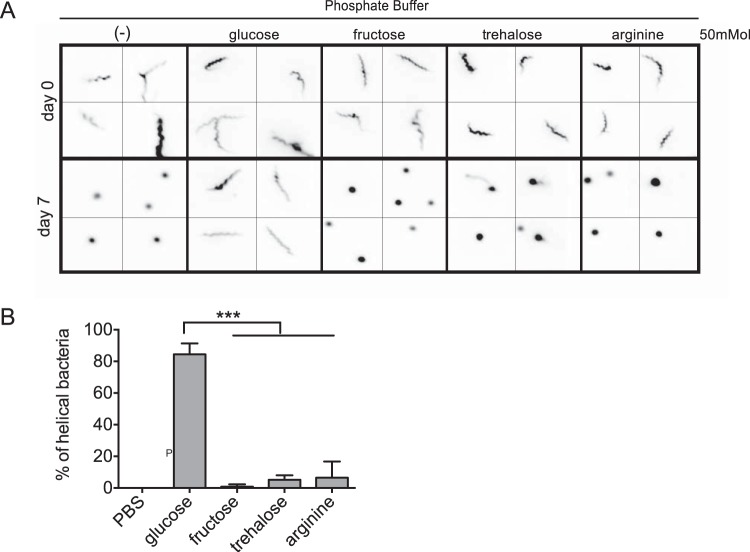
Spiroplasmas are helical and motile members of a cell wall-less eubacterial group called Mollicutes. Although all spiroplasmas are associated with arthropods, they exhibit great diversity with respect to both their modes of transmission and their effects on their hosts; ranging from horizontally transmitted pathogens and commensals to endosymbionts that are transmitted transovarially (i.e., from mother to offspring). Here we provide the first genome sequence, along with proteomic validation, of an endosymbiotic inherited Spiroplasma bacterium, the Spiroplasma poulsonii MSRO strain harbored by Drosophila melanogaster. Comparison of the genome content of S. poulsonii with that of horizontally transmitted spiroplasmas indicates that S. poulsonii has lost many metabolic pathways and transporters, demonstrating a high level of interdependence with its insect host. Consistent with genome analysis, experimental studies showed that S. poulsonii metabolizes glucose but not trehalose. Notably, trehalose is more abundant than glucose in Drosophila hemolymph, and the inability to metabolize trehalose may prevent S. poulsonii from overproliferating. Our study identifies putative virulence genes, notably, those for a chitinase, the H2O2-producing glycerol-3-phosphate oxidase, and enzymes involved in the synthesis of the eukaryote-toxic lipid cardiolipin. S. poulsonii also expresses on the cell membrane one functional adhesion-related protein and two divergent spiralin proteins that have been implicated in insect cell invasion in other spiroplasmas. These lipoproteins may be involved in the colonization of the Drosophila germ line, ensuring S. poulsonii vertical transmission. The S. poulsonii genome is a valuable resource to explore the mechanisms of male killing and symbiont-mediated protection, two cardinal features of many facultative endosymbionts.
Importance: Most insect species, including important disease vectors and crop pests, harbor vertically transmitted endosymbiotic bacteria. These endosymbionts play key roles in their hosts' fitness, including protecting them against natural enemies and manipulating their reproduction in ways that increase the frequency of symbiont infection. Little is known about the molecular mechanisms that underlie these processes. Here, we provide the first genome draft of a vertically transmitted male-killing Spiroplasma bacterium, the S. poulsonii MSRO strain harbored by D. melanogaster. Analysis of the S. poulsonii genome was complemented by proteomics and ex vivo metabolic experiments. Our results indicate that S. poulsonii has reduced metabolic capabilities and expresses divergent membrane lipoproteins and potential virulence factors that likely participate in Spiroplasma-host interactions. This work fills a gap in our knowledge of insect endosymbionts and provides tools with which to decipher the interaction between Spiroplasma bacteria and their well-characterized host D. melanogaster, which is emerging as a model of endosymbiosis.
Copyright © 2015 Paredes et al.
Figures
References
-
- Gasparich GE, Whitcomb RF, Dodge D, French FE, Glass J, Williamson DL. 2004. The genus Spiroplasma and its non-helical descendants: phylogenetic classification, correlation with phenotype and roots of the Mycoplasma mycoides clade. Int J Syst Evol Microbiol 54:893–918. doi:10.1099/ijs.0.02688-0. - DOI - PubMed
-
- Whitcomb RF, Chen TA, Williamson DL, Liao C, Tully JG, Bové JM, Mouches C, Rose DL, Coan ME, Clark TB. 1986. Spiroplasma kunkelii sp. nov.: characterization of the etiological agent of corn stunt disease. Int J Syst Bacteriol 36:170–178. doi:10.1099/00207713-36-2-170. - DOI
-
- Saillard C, Vignault JC, Bové JM, Raie A, Tully JG, Williamson DL, Fos A, Garnier M, Gadeau A, Carle P, Whitcomb RF. 1987. Spiroplasma phoeniceum sp. nov., a new plant-pathogenic species from Syria. Int J Syst Evol Microbiol 37:106–115. doi:10.1099/00207713-37-2-106. - DOI
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
