

Virus-induced dormancy in the archaeon Sulfolobus islandicus
- PMID: 25827422
- PMCID: PMC4453537
- DOI: 10.1128/mBio.02565-14
Virus-induced dormancy in the archaeon Sulfolobus islandicus
Abstract
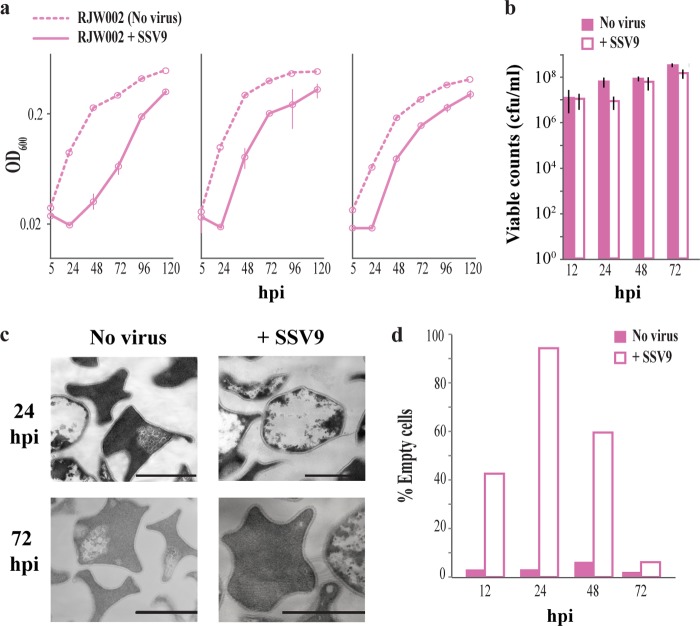
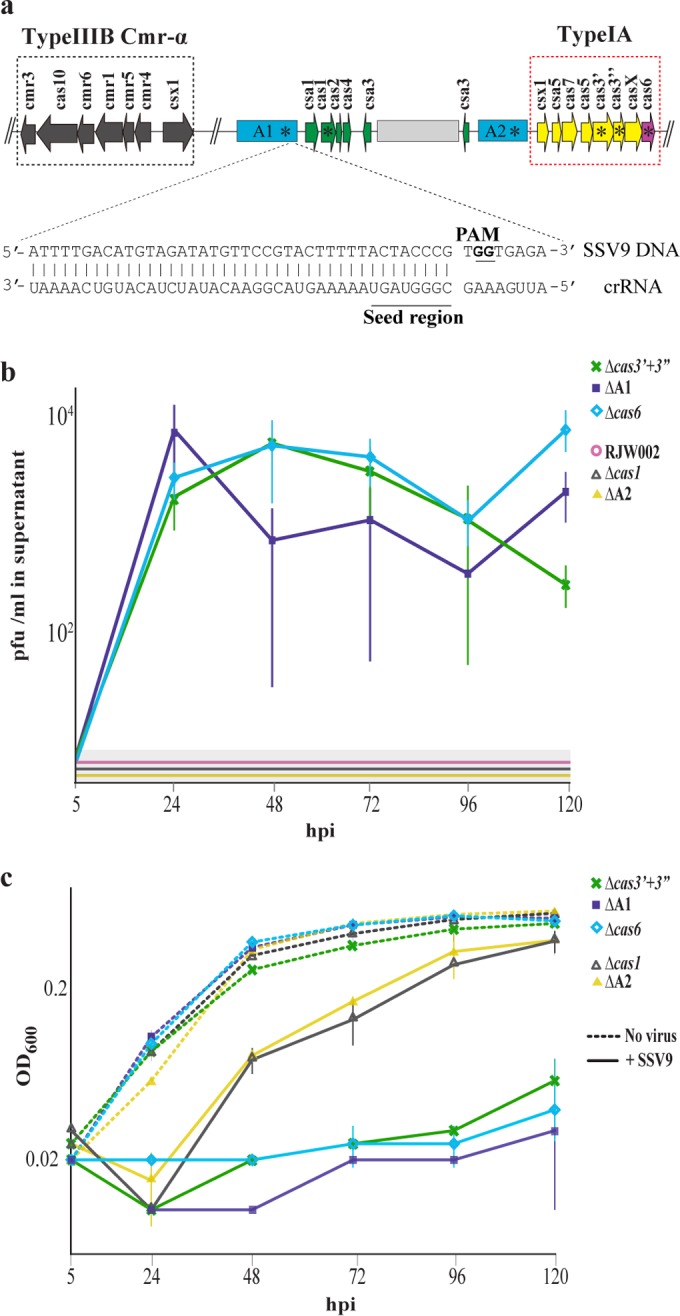
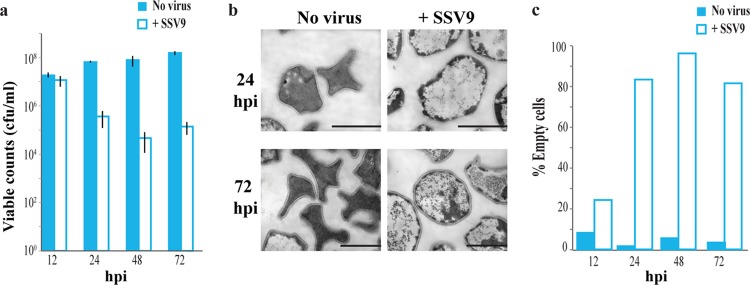
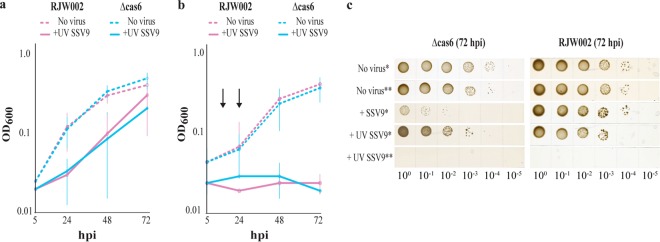
We investigated the interaction between Sulfolobus spindle-shaped virus (SSV9) and its native archaeal host Sulfolobus islandicus. We show that upon exposure to SSV9, S. islandicus strain RJW002 has a significant growth delay where the majority of cells are dormant (viable but not growing) for 24 to 48 hours postinfection (hpi) compared to the growth of controls without virus. We demonstrate that in this system, dormancy (i) is induced by both active and inactive virus particles at a low multiplicity of infection (MOI), (ii) is reversible in strains with active CRISPR-Cas immunity that prevents the establishment of productive infections, and (iii) results in dramatic and rapid host death if virus persists in the culture even at low levels. Our results add a new dimension to evolutionary models of virus-host interactions, showing that the mere presence of a virus induces host cell stasis and death independent of infection. This novel, highly sensitive, and risky bet-hedging antiviral response must be integrated into models of virus-host interactions in this system so that the true ecological impact of viruses can be predicted and understood.
Importance: Viruses of microbes play key roles in microbial ecology; however, our understanding of viral impact on host physiology is based on a few model bacteria that represent a small fraction of the life history strategies employed by hosts or viruses across the three domains that encompass the microbial world. We have demonstrated that rare and even inactive viruses induce dormancy in the model archaeon S. islandicus. Similar virus-induced dormancy strategies in other microbial systems may help to explain several confounding observations in other systems, including the surprising abundance of dormant cell types found in many microbial environments, the difficulty of culturing microorganisms in the laboratory, and the paradoxical virus-to-host abundances that do not match model predictions. A more accurate grasp of virus-host interactions will expand our understanding of the impact of viruses in microbial ecology.
Copyright © 2015 Bautista et al.
Figures
References
-
- Bohannan BJM, Lenski RE. 2000. Linking genetic change to community evolution: insights from studies of bacteria and bacteriophage. Ecol Lett 3:362–377. doi: 10.1046/j.1461-0248.2000.00161.x. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
