

No dopamine cell loss or changes in cytoskeleton function in transgenic mice expressing physiological levels of wild type or G2019S mutant LRRK2 and in human fibroblasts
- PMID: 25830304
- PMCID: PMC4382199
- DOI: 10.1371/journal.pone.0118947
No dopamine cell loss or changes in cytoskeleton function in transgenic mice expressing physiological levels of wild type or G2019S mutant LRRK2 and in human fibroblasts
Abstract
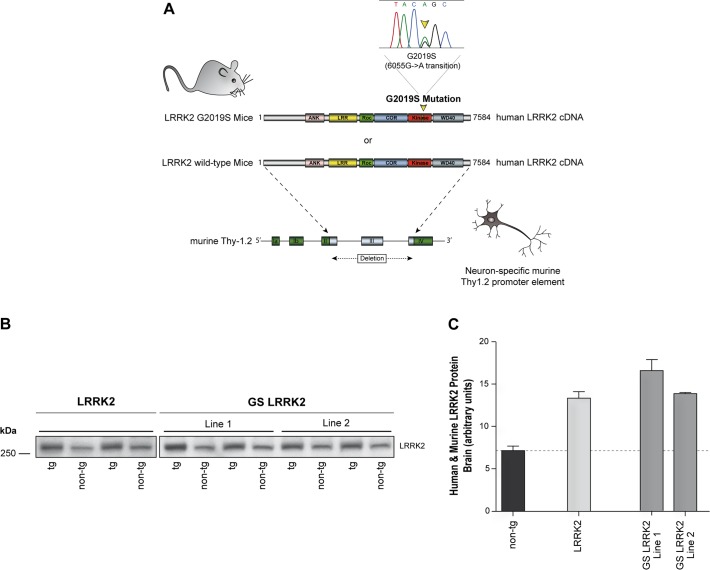
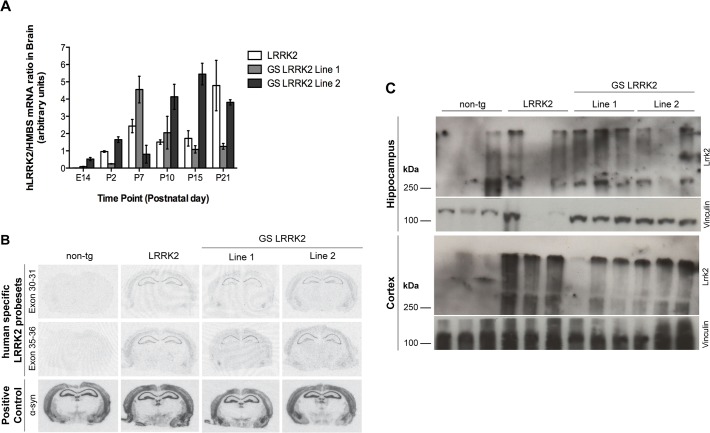
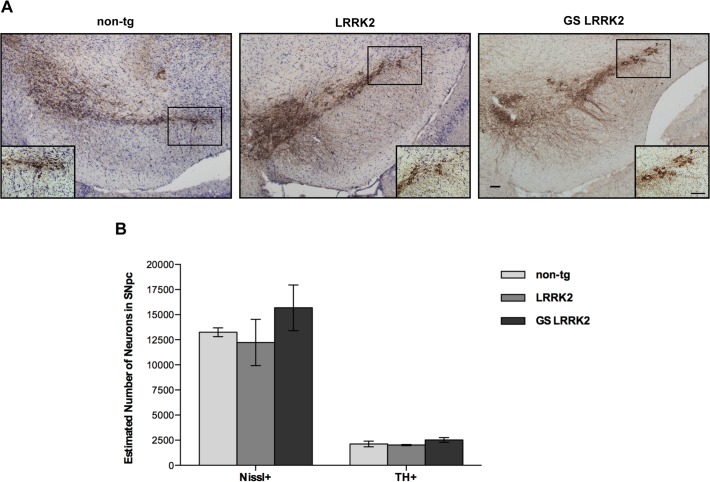
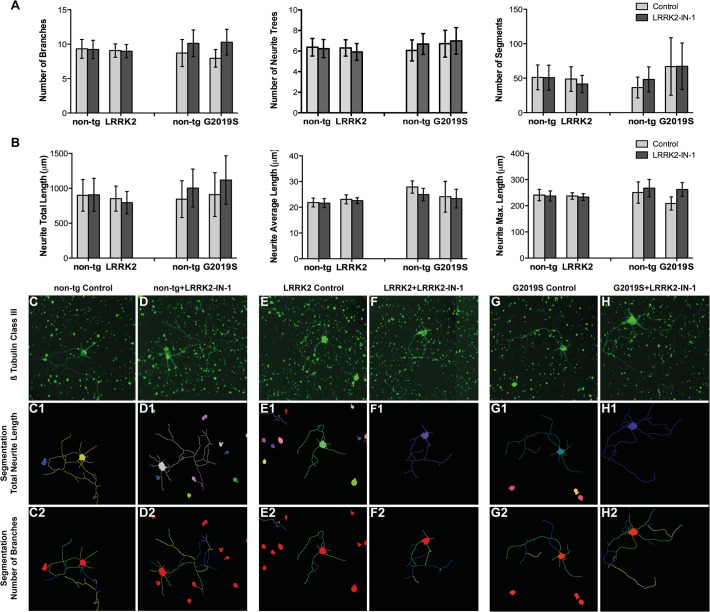
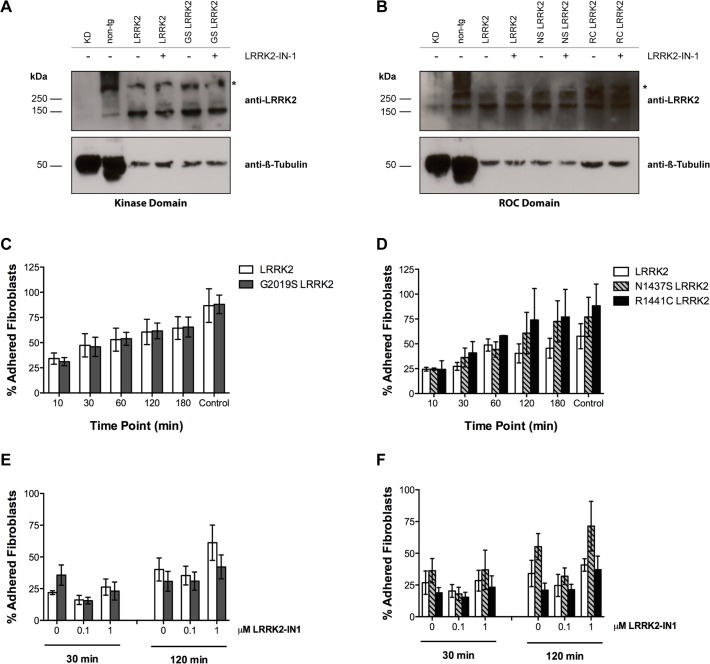
Mutations within the LRRK2 gene have been identified in Parkinson's disease (PD) patients and have been implicated in the dysfunction of several cellular pathways. Here, we explore how pathogenic mutations and the inhibition of LRRK2 kinase activity affect cytoskeleton dynamics in mouse and human cell systems. We generated and characterized a novel transgenic mouse model expressing physiological levels of human wild type and G2019S-mutant LRRK2. No neuronal loss or neurodegeneration was detected in midbrain dopamine neurons at the age of 12 months. Postnatal hippocampal neurons derived from transgenic mice showed no alterations in the seven parameters examined concerning neurite outgrowth sampled automatically on several hundred neurons using high content imaging. Treatment with the kinase inhibitor LRRK2-IN-1 resulted in no significant changes in the neurite outgrowth. In human fibroblasts we analyzed whether pathogenic LRRK2 mutations change cytoskeleton functions such as cell adhesion. To this end we compared the adhesion characteristics of human skin fibroblasts derived from six PD patients carrying one of three different pathogenic LRRK2 mutations and from four age-matched control individuals. The mutant LRRK2 variants as well as the inhibition of LRRK2 kinase activity did not reveal any significant cell adhesion differences in cultured fibroblasts. In summary, our results in both human and mouse cell systems suggest that neither the expression of wild type or mutant LRRK2, nor the inhibition of LRRK2 kinase activity affect neurite complexity and cellular adhesion.
Conflict of interest statement
Figures
Similar articles
-
14-3-3 Proteins regulate mutant LRRK2 kinase activity and neurite shortening.Hum Mol Genet. 2016 Jan 1;25(1):109-22. doi: 10.1093/hmg/ddv453. Epub 2015 Nov 5. Hum Mol Genet. 2016. PMID: 26546614 Free PMC article.
-
Selective expression of Parkinson's disease-related Leucine-rich repeat kinase 2 G2019S missense mutation in midbrain dopaminergic neurons impairs dopamine release and dopaminergic gene expression.Hum Mol Genet. 2015 Sep 15;24(18):5299-312. doi: 10.1093/hmg/ddv249. Epub 2015 Jun 29. Hum Mol Genet. 2015. PMID: 26123485 Free PMC article.
-
LRRK2 G2019S-induced mitochondrial DNA damage is LRRK2 kinase dependent and inhibition restores mtDNA integrity in Parkinson's disease.Hum Mol Genet. 2017 Nov 15;26(22):4340-4351. doi: 10.1093/hmg/ddx320. Hum Mol Genet. 2017. PMID: 28973664 Free PMC article.
-
Leucine-rich repeat kinase 2 inhibitors: a review of recent patents (2011 - 2013).Expert Opin Ther Pat. 2014 Jul;24(7):745-57. doi: 10.1517/13543776.2014.907275. Epub 2014 Jun 11. Expert Opin Ther Pat. 2014. PMID: 24918198 Review.
-
Leucine-rich repeat kinase 2 inhibitors: a patent review (2006 - 2011).Expert Opin Ther Pat. 2012 Dec;22(12):1415-26. doi: 10.1517/13543776.2012.729041. Epub 2012 Nov 6. Expert Opin Ther Pat. 2012. PMID: 23126385 Review.
Cited by
-
Pathogenic LRRK2 requires secondary factors to induce cellular toxicity.Biosci Rep. 2020 Oct 30;40(10):BSR20202225. doi: 10.1042/BSR20202225. Biosci Rep. 2020. PMID: 32975566 Free PMC article.
-
Non-cell autonomous mechanism of Parkinson's disease pathology caused by G2019S LRRK2 mutation in Ashkenazi Jewish patient: Single cell analysis.Brain Res. 2019 Nov 1;1722:146342. doi: 10.1016/j.brainres.2019.146342. Epub 2019 Jul 19. Brain Res. 2019. PMID: 31330122 Free PMC article. Review.
-
The Current State-of-the Art of LRRK2-Based Biomarker Assay Development in Parkinson's Disease.Front Neurosci. 2020 Aug 18;14:865. doi: 10.3389/fnins.2020.00865. eCollection 2020. Front Neurosci. 2020. PMID: 33013290 Free PMC article. Review.
-
Functional Impairment in Miro Degradation and Mitophagy Is a Shared Feature in Familial and Sporadic Parkinson's Disease.Cell Stem Cell. 2016 Dec 1;19(6):709-724. doi: 10.1016/j.stem.2016.08.002. Epub 2016 Sep 8. Cell Stem Cell. 2016. PMID: 27618216 Free PMC article.
-
Deletion of lrrk2 causes early developmental abnormalities and age-dependent increase of monoamine catabolism in the zebrafish brain.PLoS Genet. 2021 Sep 13;17(9):e1009794. doi: 10.1371/journal.pgen.1009794. eCollection 2021 Sep. PLoS Genet. 2021. PMID: 34516550 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
