

SWI/SNF chromatin-remodeling complexes function in noncoding RNA-dependent assembly of nuclear bodies
- PMID: 25831520
- PMCID: PMC4394320
- DOI: 10.1073/pnas.1423819112
SWI/SNF chromatin-remodeling complexes function in noncoding RNA-dependent assembly of nuclear bodies
Abstract
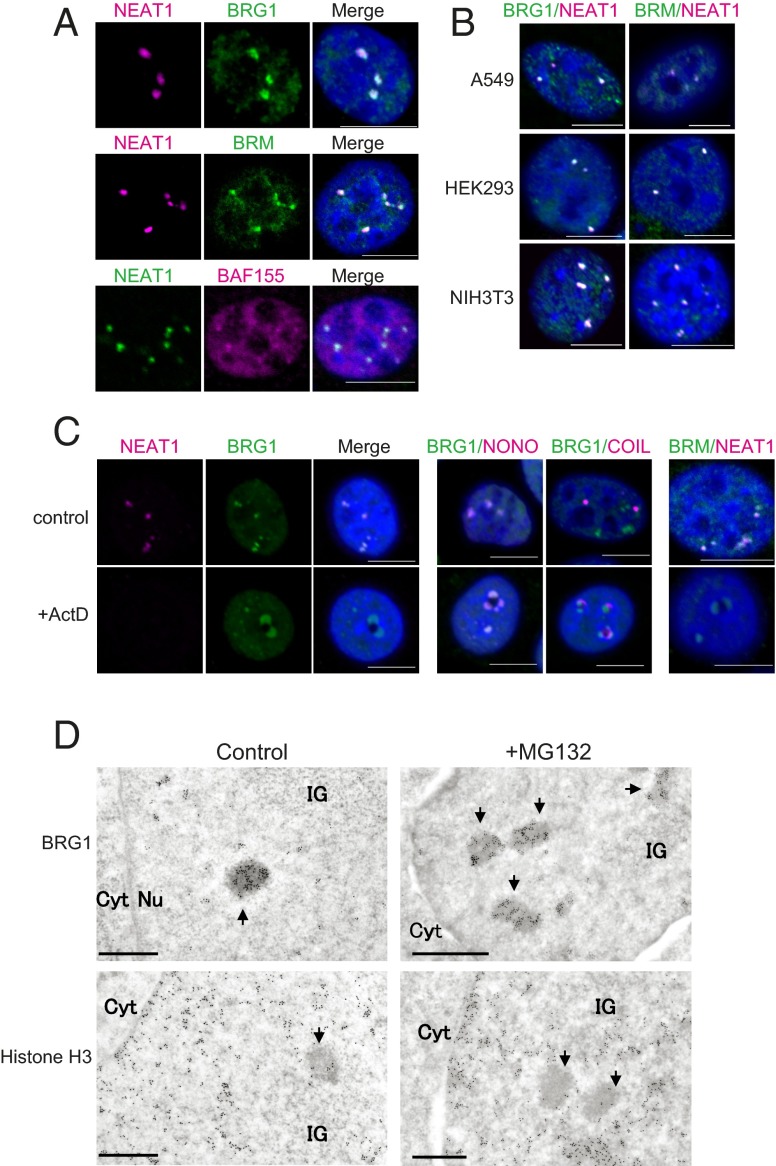
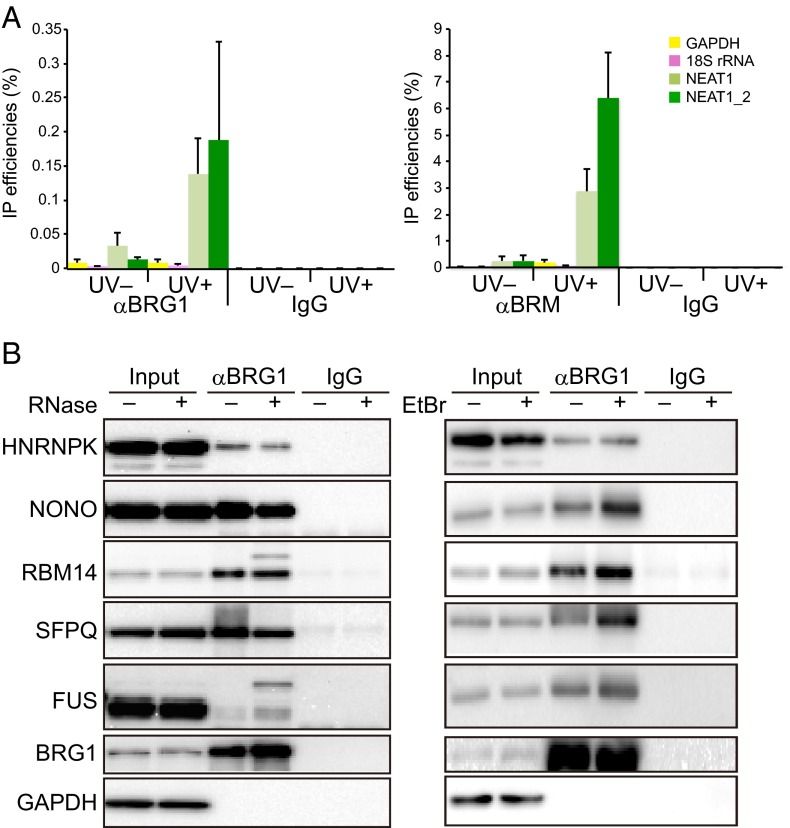
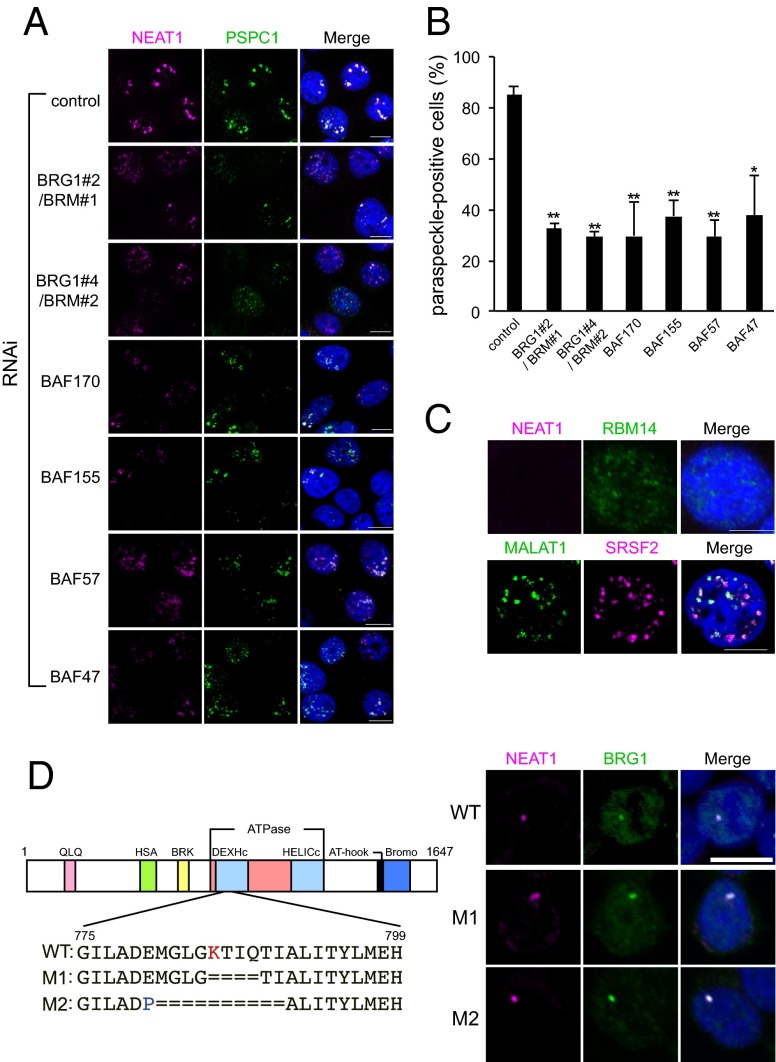
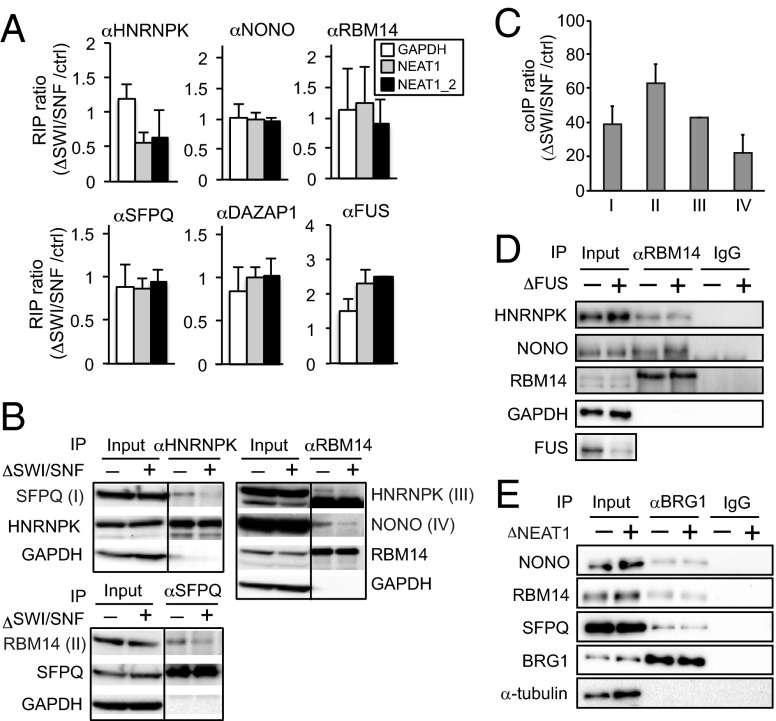
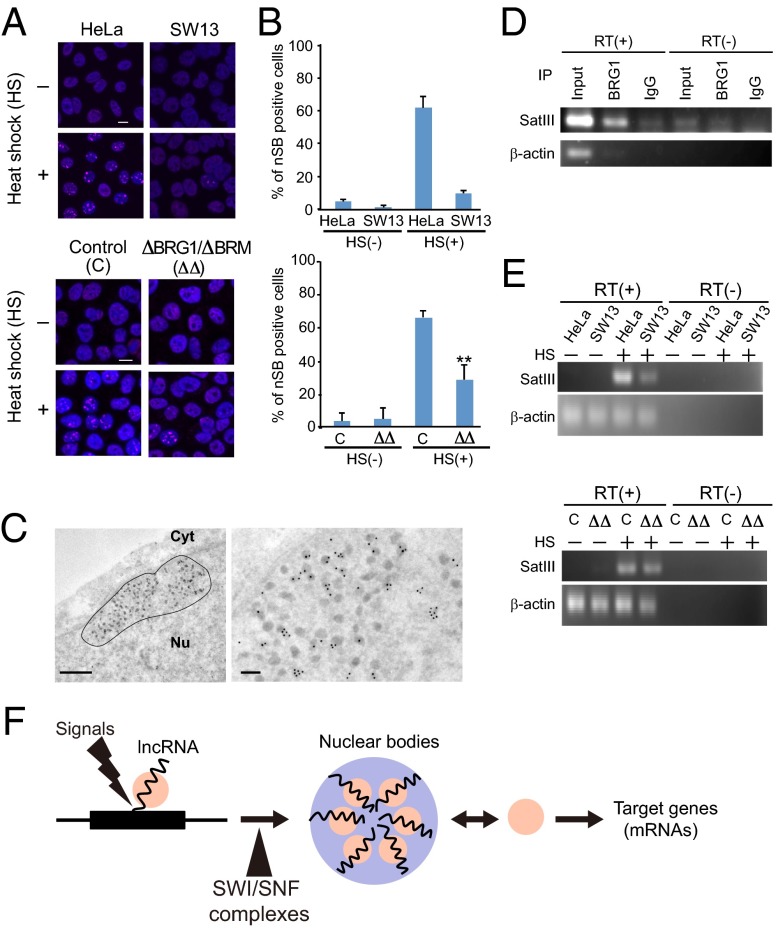
Paraspeckles are subnuclear structures that form around nuclear paraspeckle assembly transcript 1 (NEAT1) long noncoding RNA (lncRNA). Recently, paraspeckles were shown to be functional nuclear bodies involved in stress responses and the development of specific organs. Paraspeckle formation is initiated by transcription of the NEAT1 chromosomal locus and proceeds in conjunction with NEAT1 lncRNA biogenesis and a subsequent assembly step involving >40 paraspeckle proteins (PSPs). In this study, subunits of SWItch/Sucrose NonFermentable (SWI/SNF) chromatin-remodeling complexes were identified as paraspeckle components that interact with PSPs and NEAT1 lncRNA. EM observations revealed that SWI/SNF complexes were enriched in paraspeckle subdomains depleted of chromatin. Knockdown of SWI/SNF components resulted in paraspeckle disintegration, but mutation of the ATPase domain of the catalytic subunit BRG1 did not affect paraspeckle integrity, indicating that the essential role of SWI/SNF complexes in paraspeckle formation does not require their canonical activity. Knockdown of SWI/SNF complexes barely affected the levels of known essential paraspeckle components, but markedly diminished the interactions between essential PSPs, suggesting that SWI/SNF complexes facilitate organization of the PSP interaction network required for intact paraspeckle assembly. The interactions between SWI/SNF components and essential PSPs were maintained in NEAT1-depleted cells, suggesting that SWI/SNF complexes not only facilitate interactions between PSPs, but also recruit PSPs during paraspeckle assembly. SWI/SNF complexes were also required for Satellite III lncRNA-dependent formation of nuclear stress bodies under heat-shock conditions. Our data suggest the existence of a common mechanism underlying the formation of lncRNA-dependent nuclear body architectures in mammalian cells.
Keywords: chromatin-remodeling complex; long noncoding RNA; nuclear bodies; ribonucleoprotein assembly.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Chromatin remodeling complexes in the assembly of long noncoding RNA-dependent nuclear bodies.Nucleus. 2015;6(6):462-7. doi: 10.1080/19491034.2015.1119353. Epub 2015 Dec 28. Nucleus. 2015. PMID: 26709446 Free PMC article.
-
Molecular anatomy of the architectural NEAT1 noncoding RNA: The domains, interactors, and biogenesis pathway required to build phase-separated nuclear paraspeckles.Wiley Interdiscip Rev RNA. 2019 Nov;10(6):e1545. doi: 10.1002/wrna.1545. Epub 2019 May 1. Wiley Interdiscip Rev RNA. 2019. PMID: 31044562 Review.
-
Paraspeckle formation during the biogenesis of long non-coding RNAs.RNA Biol. 2013 Mar;10(3):456-61. doi: 10.4161/rna.23547. Epub 2013 Jan 16. RNA Biol. 2013. PMID: 23324609 Free PMC article. Review.
-
Highly ordered spatial organization of the structural long noncoding NEAT1 RNAs within paraspeckle nuclear bodies.Mol Biol Cell. 2010 Nov 15;21(22):4020-7. doi: 10.1091/mbc.E10-08-0690. Epub 2010 Sep 29. Mol Biol Cell. 2010. PMID: 20881053 Free PMC article.
-
Organization and function of paraspeckles.Essays Biochem. 2020 Dec 7;64(6):875-882. doi: 10.1042/EBC20200010. Essays Biochem. 2020. PMID: 32830222 Review.
Cited by
-
Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer.Nat Genet. 2016 May;48(5):500-9. doi: 10.1038/ng.3547. Epub 2016 Apr 11. Nat Genet. 2016. PMID: 27064257
-
Unconventional roles of chromatin remodelers and long non-coding RNAs in cell division.Cell Mol Life Sci. 2023 Nov 20;80(12):365. doi: 10.1007/s00018-023-04949-8. Cell Mol Life Sci. 2023. PMID: 37982870 Free PMC article. Review.
-
Satellite RNAs: emerging players in subnuclear architecture and gene regulation.EMBO J. 2023 Sep 18;42(18):e114331. doi: 10.15252/embj.2023114331. Epub 2023 Aug 1. EMBO J. 2023. PMID: 37526230 Free PMC article. Review.
-
Nascent ribosomal RNA act as surfactant that suppresses growth of fibrillar centers in nucleolus.Commun Biol. 2023 Nov 7;6(1):1129. doi: 10.1038/s42003-023-05519-1. Commun Biol. 2023. PMID: 37935838 Free PMC article.
-
On the functional relevance of spatiotemporally-specific patterns of experience-dependent long noncoding RNA expression in the brain.RNA Biol. 2021 Jul;18(7):1025-1036. doi: 10.1080/15476286.2020.1868165. Epub 2021 Jan 4. RNA Biol. 2021. PMID: 33397182 Free PMC article. Review.
References
-
- Visa N, Puvion-Dutilleul F, Bachellerie JP, Puvion E. Intranuclear distribution of U1 and U2 snRNAs visualized by high resolution in situ hybridization: Revelation of a novel compartment containing U1 but not U2 snRNA in HeLa cells. Eur J Cell Biol. 1993;60(2):308–321. - PubMed
-
- Fox AH, et al. Paraspeckles: A novel nuclear domain. Curr Biol. 2002;12(1):13–25. - PubMed
-
- Prasanth KV, et al. Regulating gene expression through RNA nuclear retention. Cell. 2005;123(2):249–263. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
