

Emergence of hematopoietic stem and progenitor cells involves a Chd1-dependent increase in total nascent transcription
- PMID: 25831528
- PMCID: PMC4394284
- DOI: 10.1073/pnas.1424850112
Emergence of hematopoietic stem and progenitor cells involves a Chd1-dependent increase in total nascent transcription
Abstract
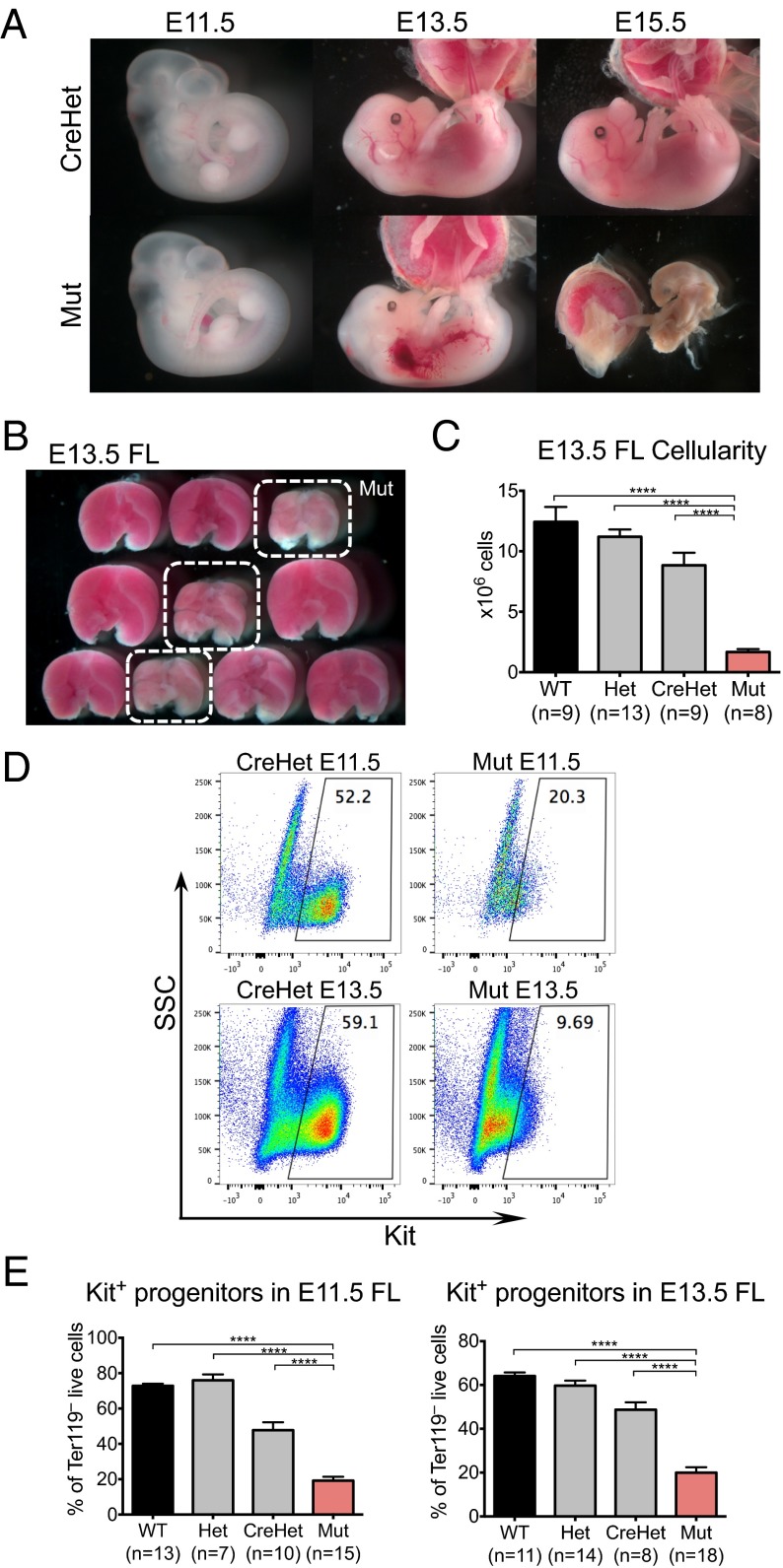
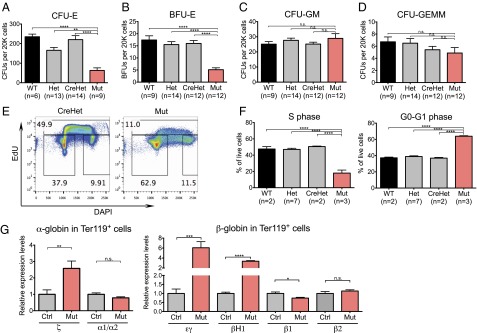
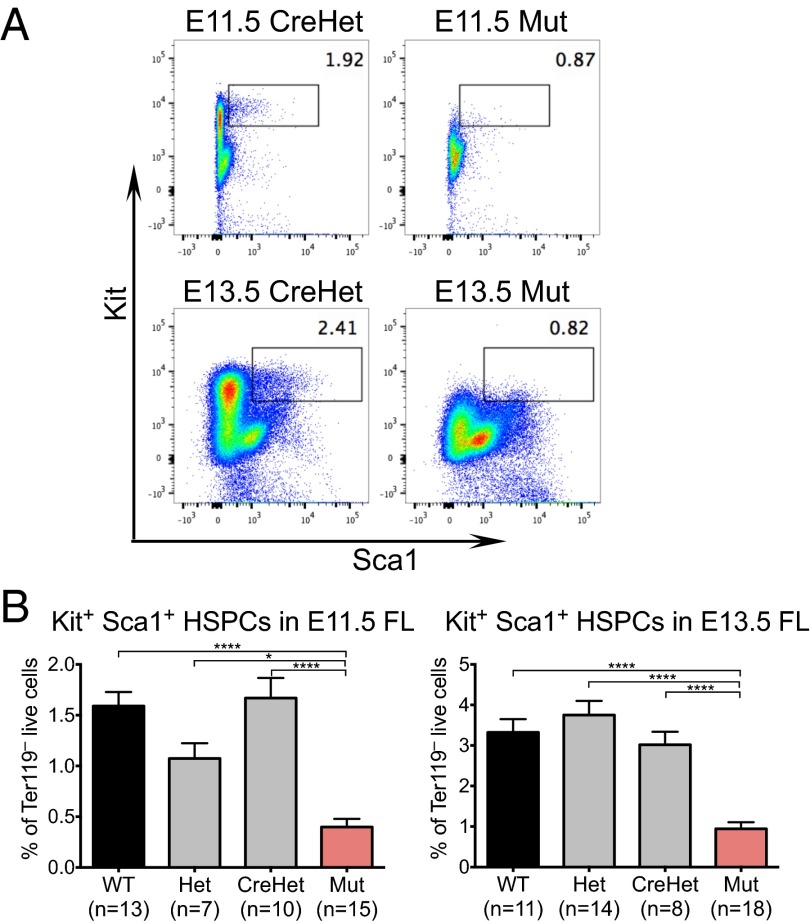
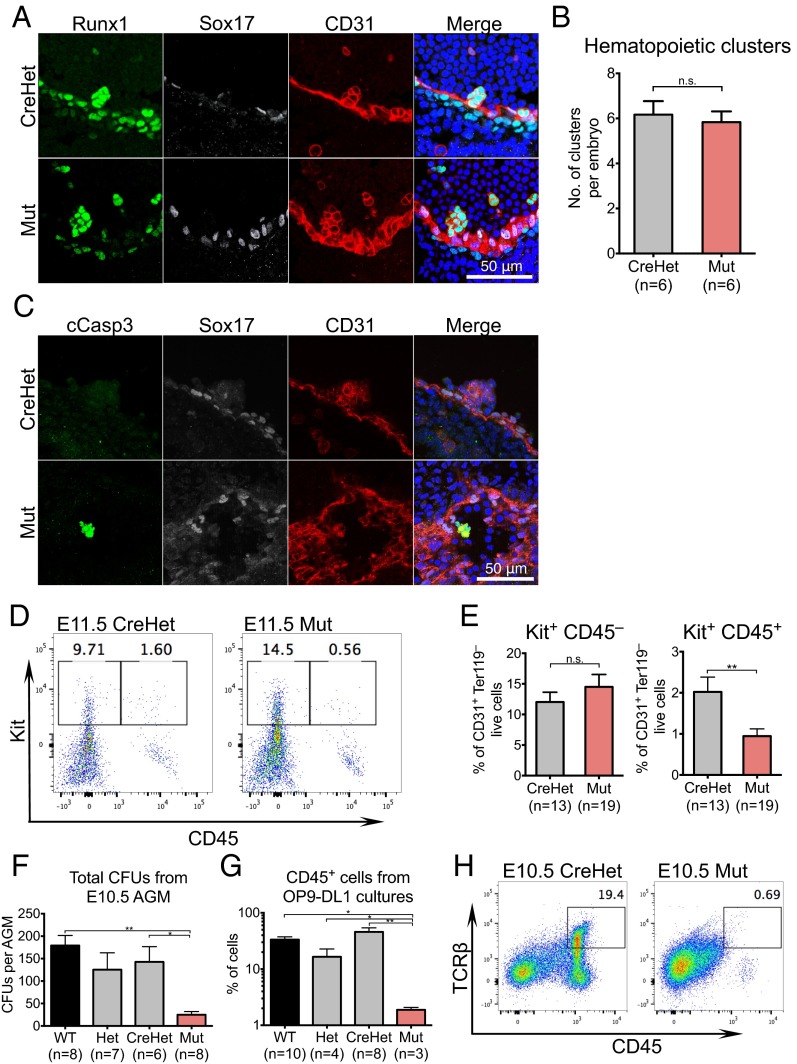
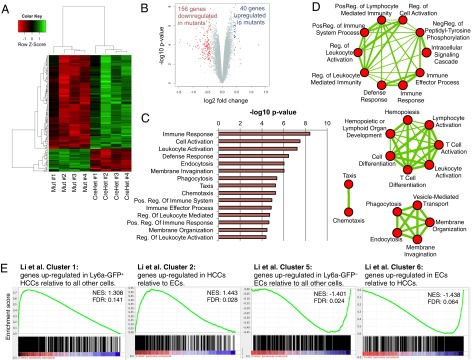
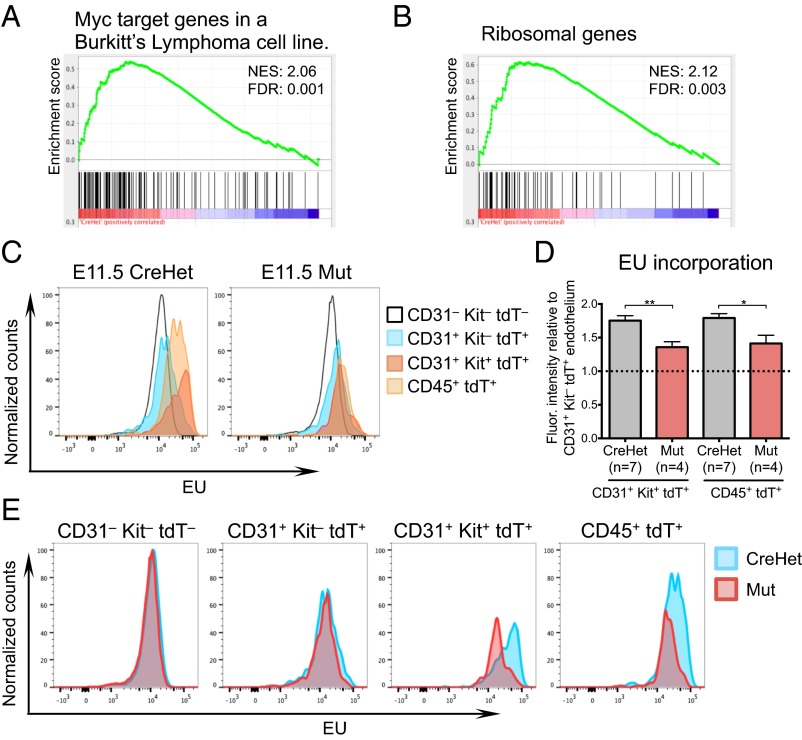
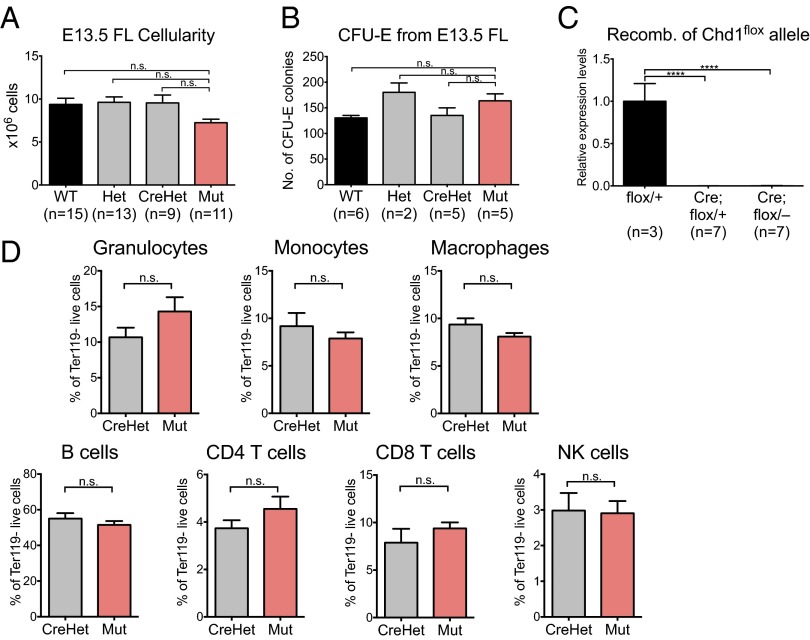
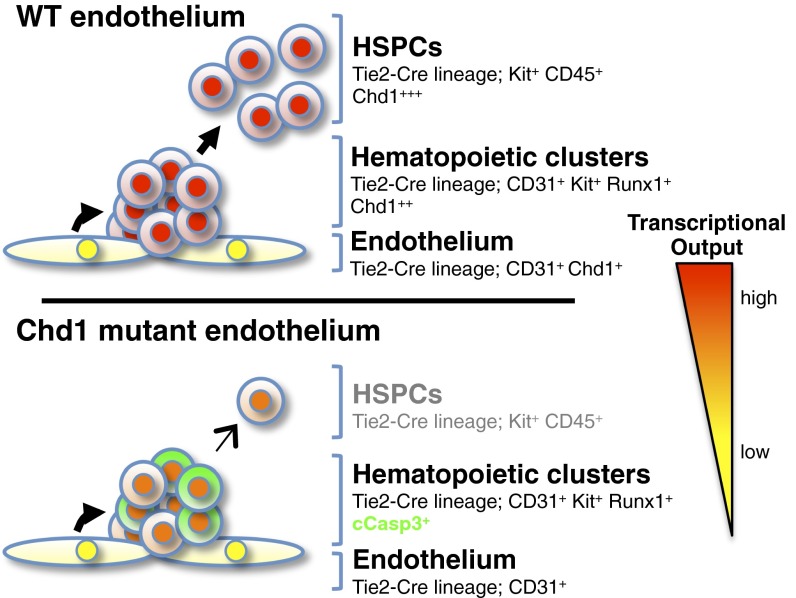
Lineage specification during development involves reprogramming of transcriptional states, but little is known about how this is regulated in vivo. The chromatin remodeler chomodomain helicase DNA-binding protein 1 (Chd1) promotes an elevated transcriptional output in mouse embryonic stem cells. Here we report that endothelial-specific deletion of Chd1 leads to loss of definitive hematopoietic progenitors, anemia, and lethality by embryonic day (E)15.5. Mutant embryos contain normal numbers of E10.5 intraaortic hematopoietic clusters that express Runx1 and Kit, but these clusters undergo apoptosis and fail to mature into blood lineages in vivo and in vitro. Hematopoietic progenitors emerging from the aorta have an elevated transcriptional output relative to structural endothelium, and this elevation is Chd1-dependent. In contrast, hematopoietic-specific deletion of Chd1 using Vav-Cre has no apparent phenotype. Our results reveal a new paradigm of regulation of a developmental transition by elevation of global transcriptional output that is critical for hemogenesis and may play roles in other contexts.
Keywords: Chd1; definitive hematopoiesis; endothelial-to-hematopoietic transition; hemogenic endothelium; transcription.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
RBPjkappa-dependent Notch function regulates Gata2 and is essential for the formation of intra-embryonic hematopoietic cells.Development. 2005 Mar;132(5):1117-26. doi: 10.1242/dev.01660. Epub 2005 Feb 2. Development. 2005. PMID: 15689374
-
Chd1 is essential for the high transcriptional output and rapid growth of the mouse epiblast.Development. 2015 Jan 1;142(1):118-27. doi: 10.1242/dev.114843. Epub 2014 Dec 5. Development. 2015. PMID: 25480920 Free PMC article.
-
The homeodomain protein Meis1 is essential for definitive hematopoiesis and vascular patterning in the mouse embryo.Dev Biol. 2005 Apr 15;280(2):307-20. doi: 10.1016/j.ydbio.2005.01.004. Dev Biol. 2005. PMID: 15882575
-
Enforced activation of STAT5A facilitates the generation of embryonic stem-derived hematopoietic stem cells that contribute to hematopoiesis in vivo.Stem Cells. 2004;22(7):1191-204. doi: 10.1634/stemcells.2004-0033. Stem Cells. 2004. PMID: 15579639
-
CHD1, a multifaceted epigenetic remodeler in prostate cancer.Front Oncol. 2023 Jan 26;13:1123362. doi: 10.3389/fonc.2023.1123362. eCollection 2023. Front Oncol. 2023. PMID: 36776288 Free PMC article. Review.
Cited by
-
The role of chromodomain helicase DNA binding protein 1 (CHD1) in promoting an invasive prostate cancer phenotype.Ther Adv Urol. 2021 Aug 4;13:17562872211022462. doi: 10.1177/17562872211022462. eCollection 2021 Jan-Dec. Ther Adv Urol. 2021. PMID: 34408788 Free PMC article.
-
Chromatin remodeling in tissue stem cell fate determination.Cell Regen. 2024 Sep 30;13(1):18. doi: 10.1186/s13619-024-00203-z. Cell Regen. 2024. PMID: 39348027 Free PMC article. Review.
-
Epigenetic and Epitranscriptomic Factors Make a Mark on Hematopoietic Stem Cell Development.Curr Stem Cell Rep. 2018 Mar;4(1):22-32. Epub 2018 Feb 3. Curr Stem Cell Rep. 2018. PMID: 29910999 Free PMC article.
-
Chd1 protects genome integrity at promoters to sustain hypertranscription in embryonic stem cells.Nat Commun. 2021 Aug 11;12(1):4859. doi: 10.1038/s41467-021-25088-3. Nat Commun. 2021. PMID: 34381042 Free PMC article.
-
Global Hypertranscription in the Mouse Embryonic Germline.Cell Rep. 2017 Jun 6;19(10):1987-1996. doi: 10.1016/j.celrep.2017.05.036. Cell Rep. 2017. PMID: 28591571 Free PMC article.
References
-
- Cumano A, Godin I. Ontogeny of the hematopoietic system. Annu Rev Immunol. 2007;25:745–785. - PubMed
-
- Palis J, Robertson S, Kennedy M, Wall C, Keller G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development. 1999;126(22):5073–5084. - PubMed
-
- de Bruijn MFTR, et al. Hematopoietic stem cells localize to the endothelial cell layer in the midgestation mouse aorta. Immunity. 2002;16(5):673–683. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
