

The Arp2/3 complex has essential roles in vesicle trafficking and transcytosis in the mammalian small intestine
- PMID: 25833710
- PMCID: PMC4472011
- DOI: 10.1091/mbc.E14-10-1481
The Arp2/3 complex has essential roles in vesicle trafficking and transcytosis in the mammalian small intestine
Abstract
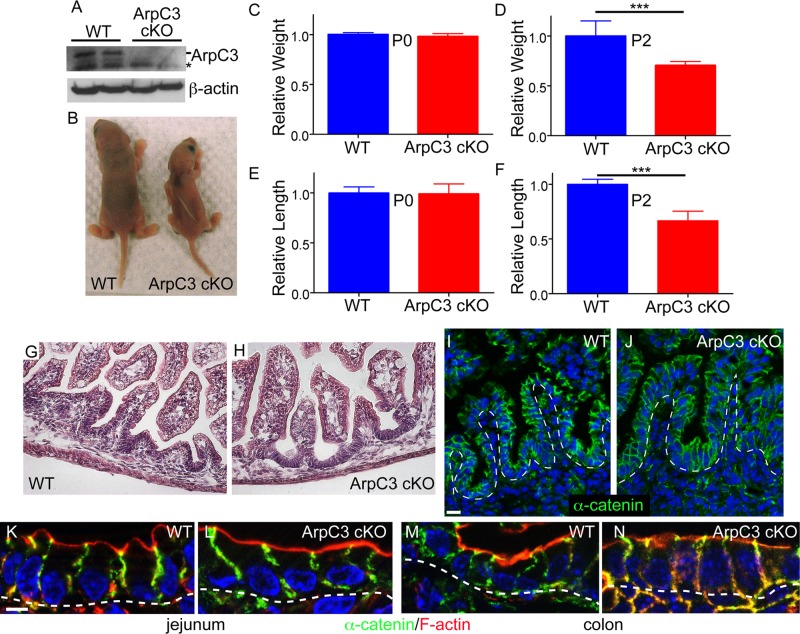
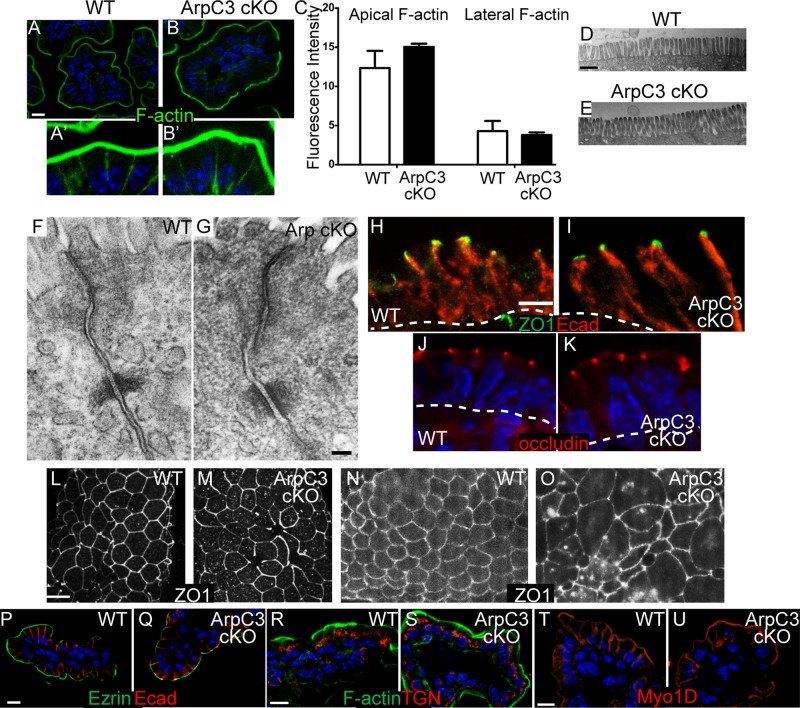
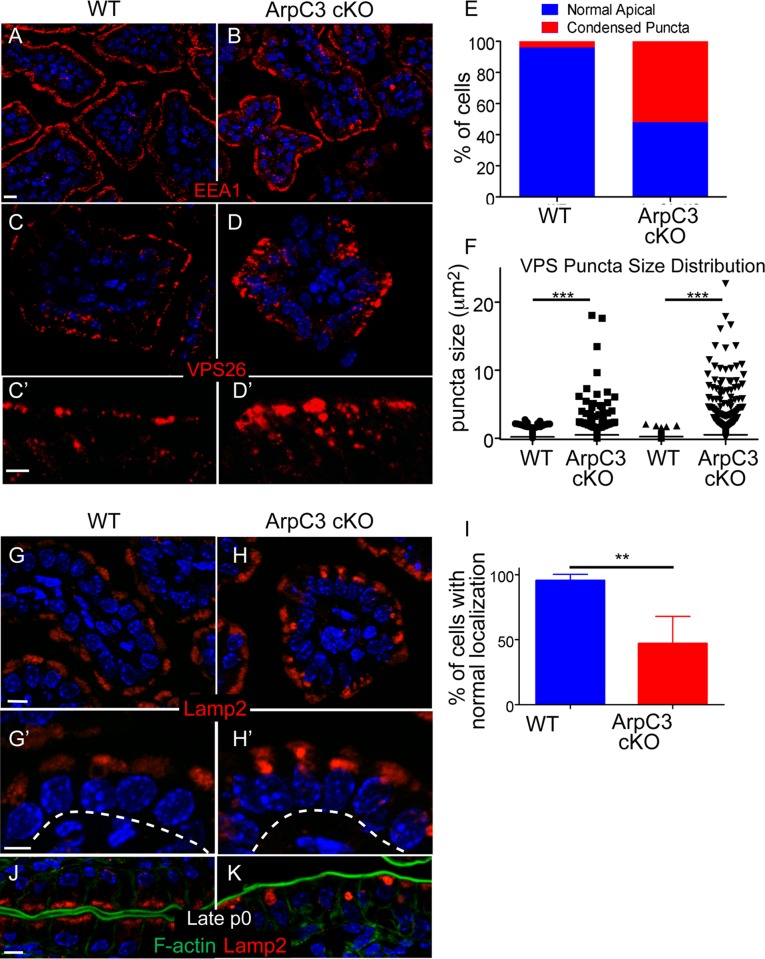
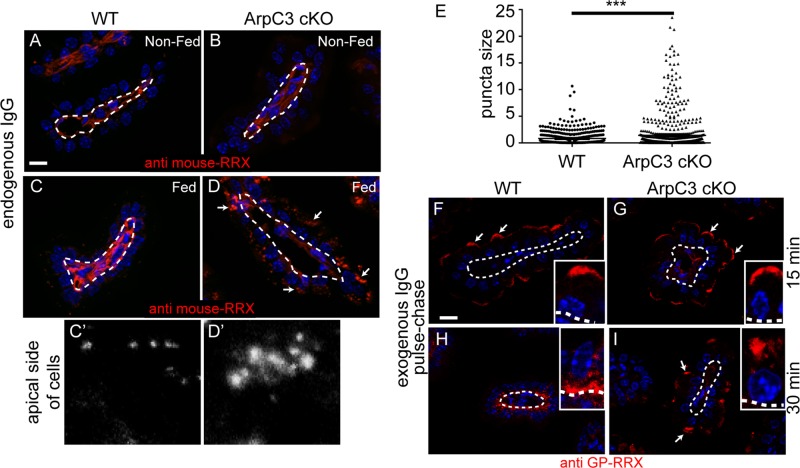
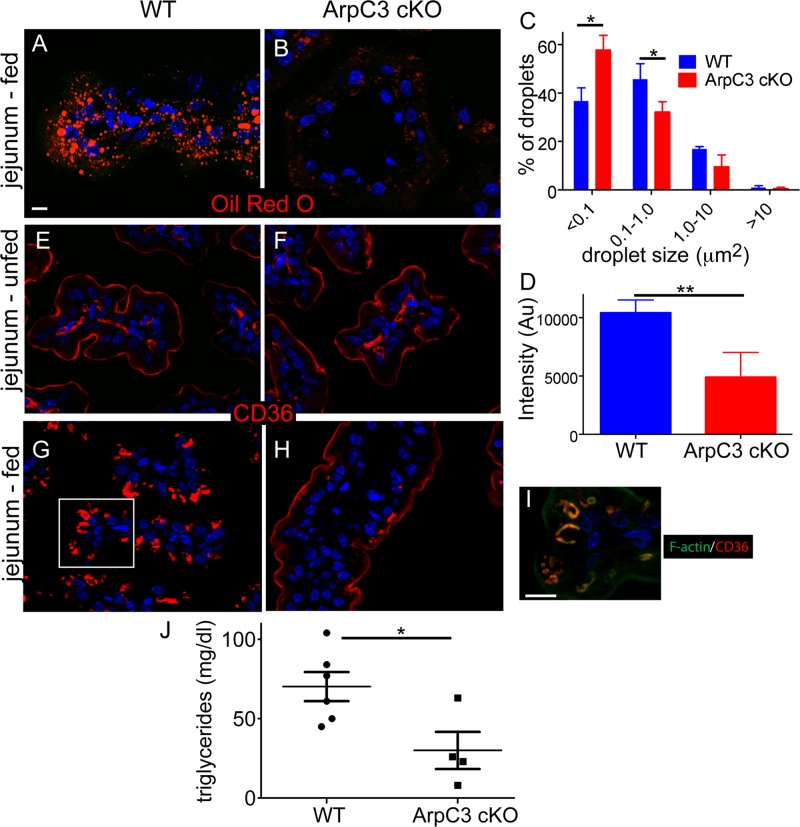
The Arp2/3 complex is the only known nucleator of branched F-actin filaments. Work in cultured cells has established a wide array of functions for this complex in controlling cell migration, shape, and adhesion. However, loss of Arp2/3 complex function in tissues has yielded cell type-specific phenotypes. Here we report essential functions of the Arp2/3 complex in the intestinal epithelium. The Arp2/3 complex was dispensable for intestinal development, generation of cortical F-actin, and cell polarity. However, it played essential roles in vesicle trafficking. We found that in the absence of ArpC3, enterocytes had defects in the organization of the endolysosomal system. These defects were physiologically relevant, as transcytosis of IgG was disrupted, lipid absorption was perturbed, and neonatal mice died within days of birth. These data highlight the important roles of the Arp2/3 complex in vesicle trafficking in enterocytes and suggest that defects in cytoplasmic F-actin assembly by the Arp2/3 complex, rather than cortical pools, underlie many of the phenotypes seen in the mutant small intestine.
© 2015 Zhou et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Chevrot M, Martin C, Passilly-Degrace P, Besnard P. Role of CD36 in oral and postoral sensing of lipids. Handbk Exp Pharmacol. 2012;2012:295–307. - PubMed
-
- Derivery E, Sousa C, Gautier JJ, Lombard B, Loew D, Gautreau A. The Arp2/3 activator WASH controls the fission of endosomes through a large multiprotein complex. Dev Cell. 2009;17:712–723. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
