

Animal models and the molecular biology of hepadnavirus infection
- PMID: 25833941
- PMCID: PMC4382722
- DOI: 10.1101/cshperspect.a021352
Animal models and the molecular biology of hepadnavirus infection
Abstract
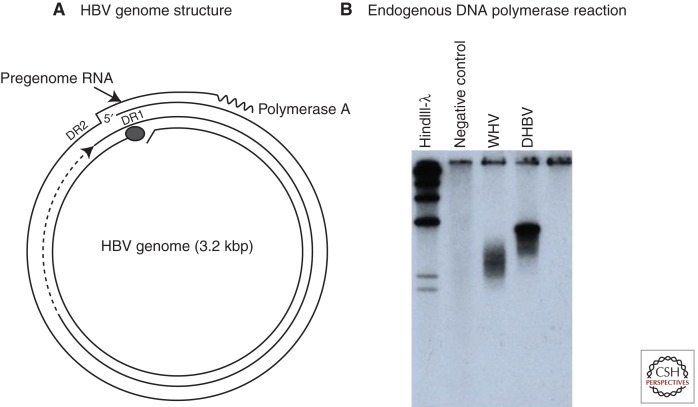
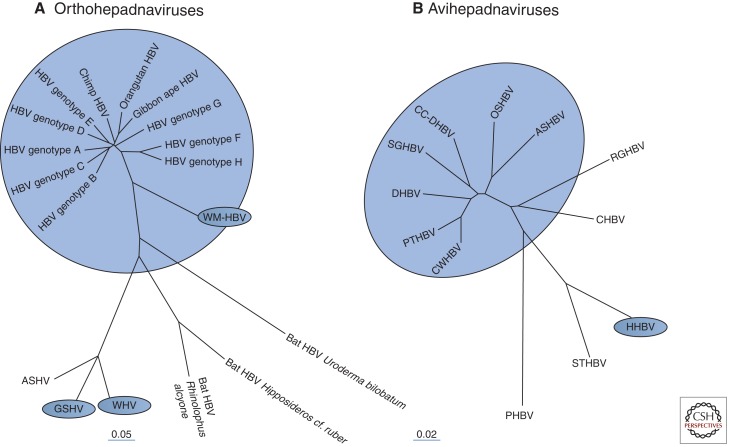
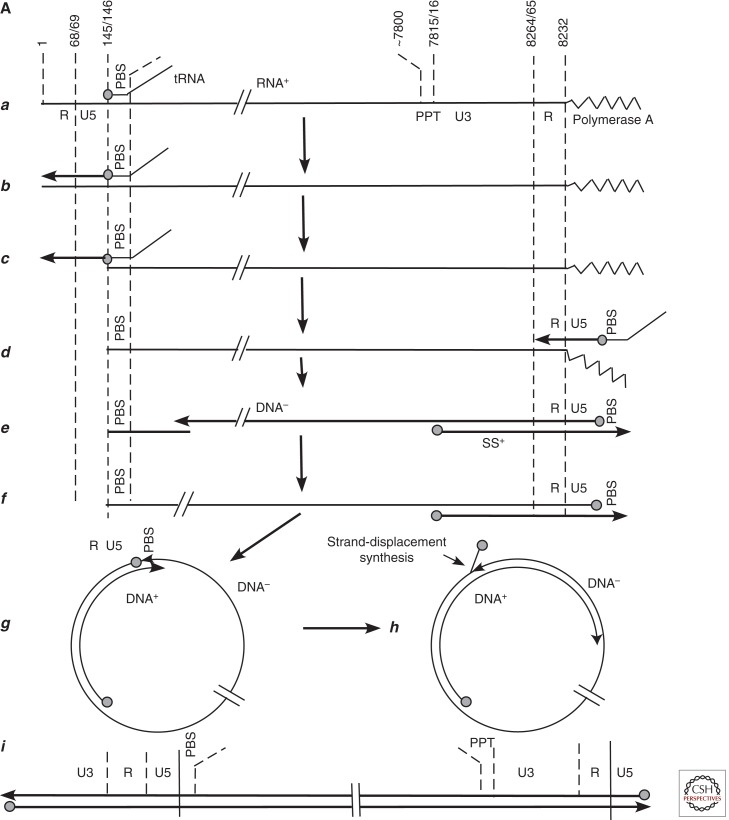
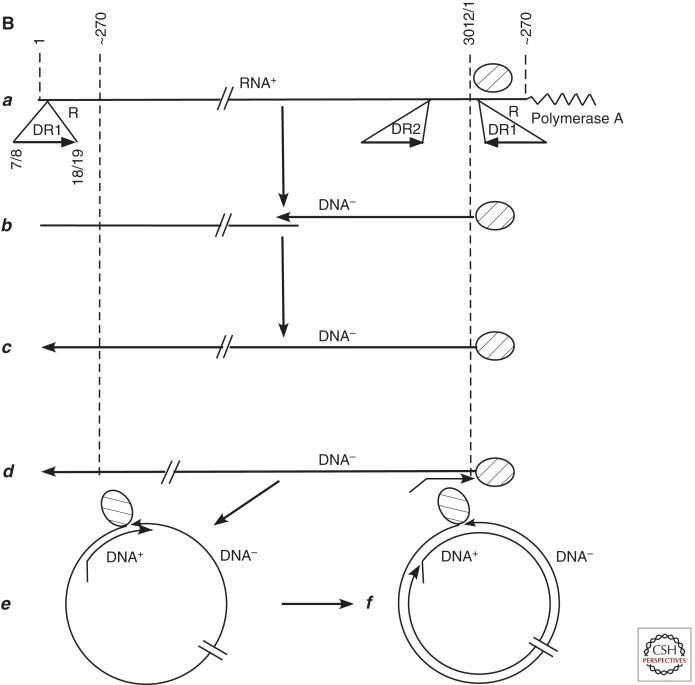
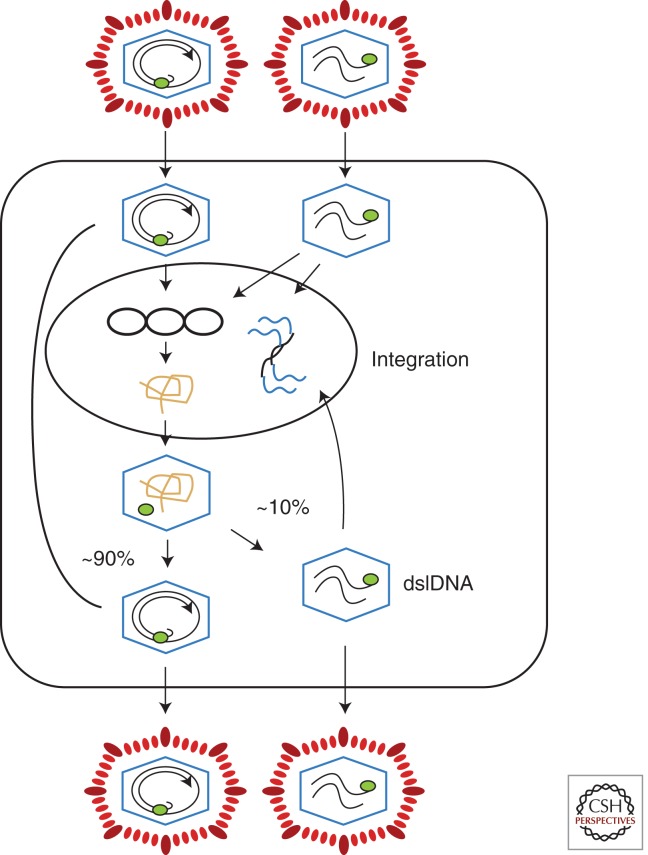
Australian antigen, the envelope protein of hepatitis B virus (HBV), was discovered in 1967 as a prevalent serum antigen in hepatitis B patients. Early electron microscopy (EM) studies showed that this antigen was present in 22-nm particles in patient sera, which were believed to be incomplete virus. Complete virus, much less abundant than the 22-nm particles, was finally visualized in 1970. HBV was soon found to infect chimpanzees, gorillas, orangutans, gibbon apes, and, more recently, tree shrews (Tupaia belangeri) and cynomolgus macaques (Macaca fascicularis). This restricted host range placed limits on the kinds of studies that might be performed to better understand the biology and molecular biology of HBV and to develop antiviral therapies to treat chronic infections. About 10 years after the discovery of HBV, this problem was bypassed with the discovery of viruses related to HBV in woodchucks, ground squirrels, and ducks. Although unlikely animal models, their use revealed the key steps in hepadnavirus replication and in the host response to infection, including the fact that the viral nuclear episome is the ultimate target for immune clearance of transient infections and antiviral therapy of chronic infections. Studies with these and other animal models have also suggested interesting clues into the link between chronic HBV infection and hepatocellular carcinoma.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Aden DP, Fogel A, Plotkin S, Damjanov I, Knowles BB 1979. Controlled synthesis of HBsAg in a differentiated human liver carcinoma-derived cell line. Nature 282: 615–616. - PubMed
-
- Alison MR, Islam S, Lim S 2009. Stem cells in liver regeneration, fibrosis and cancer: The good, the bad and the ugly. J Pathol 217: 282–298. - PubMed
-
- Bayer ME, Blumberg BS, Werner B 1968. Particles associated with Australia antigen in the sera of patients with leukaemia, Down’s syndrome and hepatitis. Nature 218: 1057–1059. - PubMed
-
- Beasley RP, Lin CC, Hwang LY, Chien CS 1981. Hepatocellular carcinoma and hepatitis B virus. Lancet 2: 1129–1133. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources