

Shake, rattle, and roll: Impact of the dynamics of flavivirus particles on their interactions with the host
- PMID: 25835729
- PMCID: PMC4900690
- DOI: 10.1016/j.virol.2015.03.025
Shake, rattle, and roll: Impact of the dynamics of flavivirus particles on their interactions with the host
Abstract
Remarkable progress in structural biology has equipped virologists with insight into structures of viral proteins and virions at increasingly high resolution. Structural information has been used extensively to address fundamental questions about virtually all aspects of how viruses replicate in cells, interact with the host, and in the design of antiviral compounds. However, many critical aspects of virology exist outside the snapshots captured by traditional methods used to generate high-resolution structures. Like all proteins, viral proteins are not static structures. The conformational flexibility and dynamics of proteins play a significant role in protein-protein interactions, and in the structure and biology of virus particles. This review will discuss the implications of the dynamics of viral proteins on the biology, antigenicity, and immunogenicity of flaviviruses.
Keywords: Antibody-mediated neutralization; Dengue virus; Flavivirus; Structural dynamics; Viral breathing; West Nile virus.
Published by Elsevier Inc.
Figures
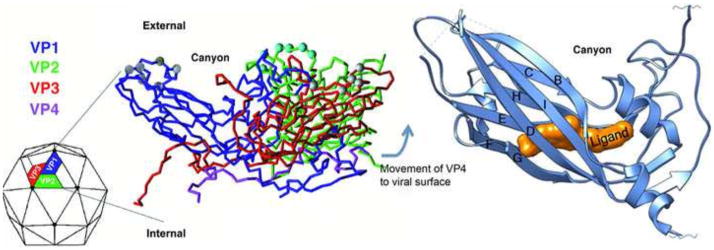
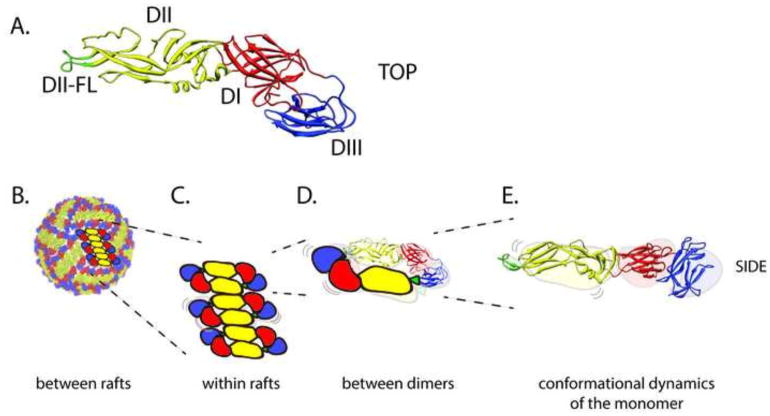
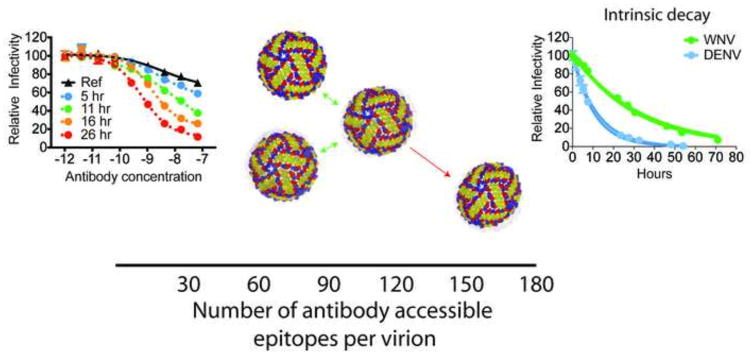
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
