

Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity-related genes, revealing gene gain and lose dynamics in genome evolution
- PMID: 25837042
- PMCID: PMC4383609
- DOI: 10.1371/journal.ppat.1004801
Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity-related genes, revealing gene gain and lose dynamics in genome evolution
Abstract
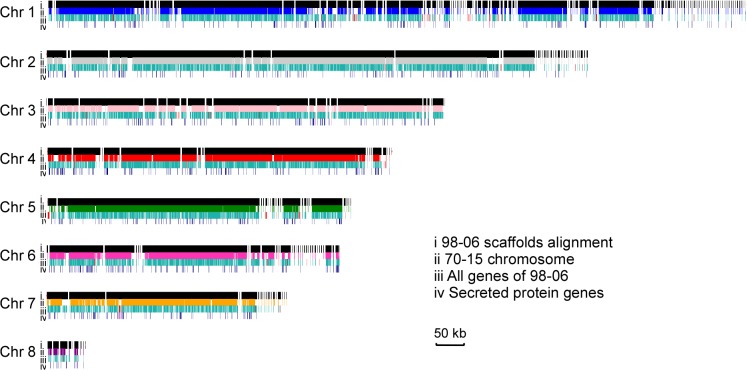
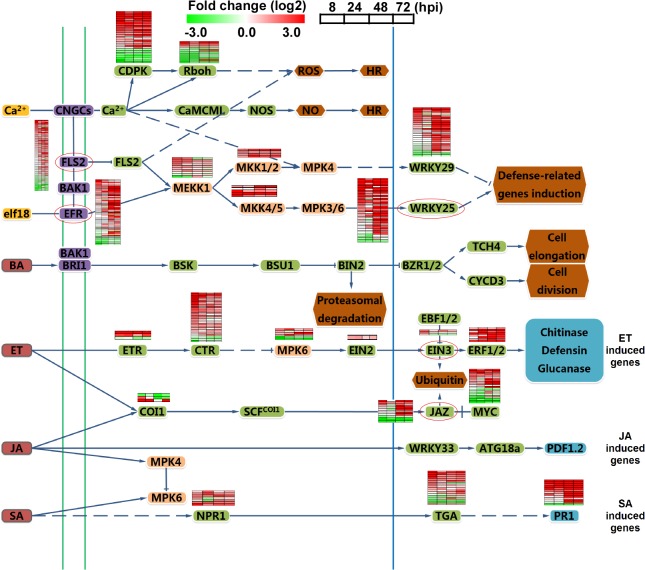
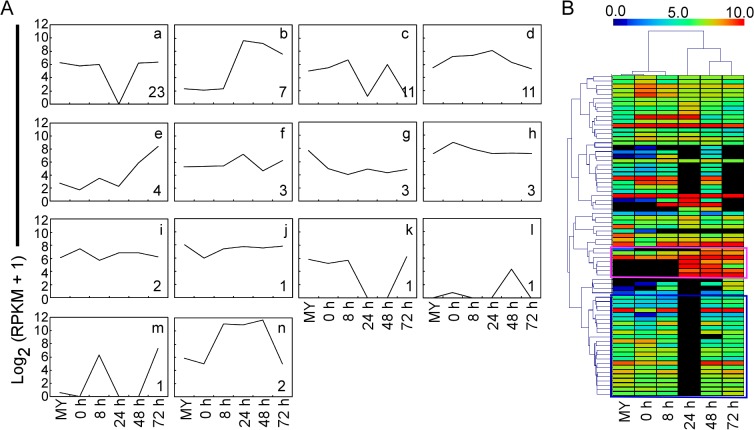
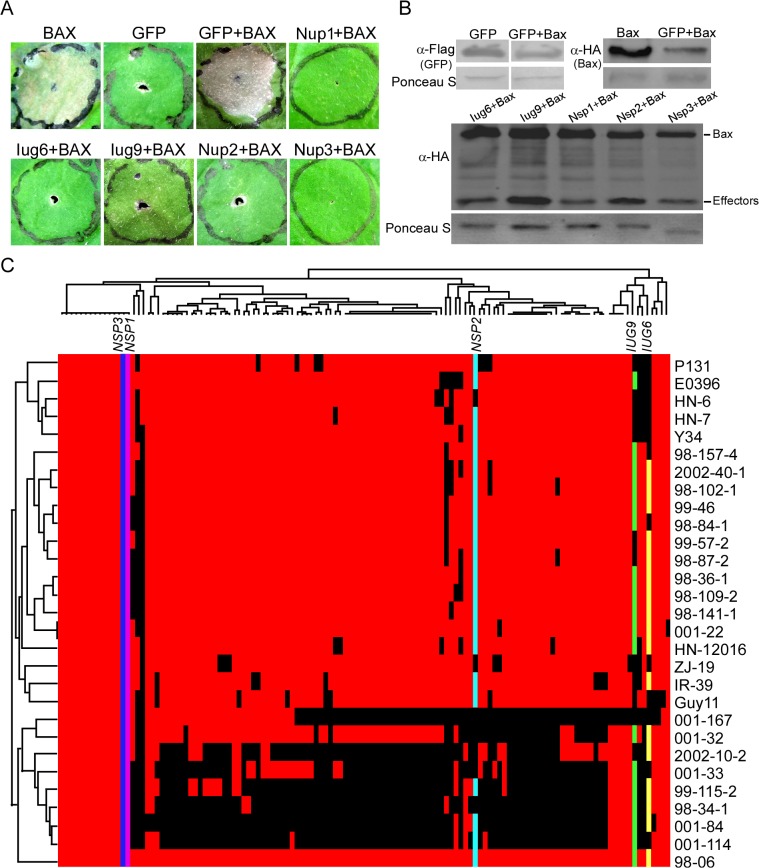
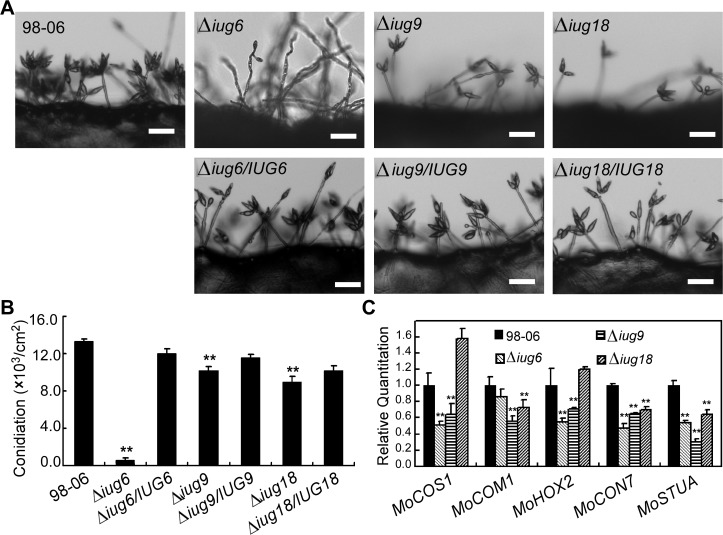
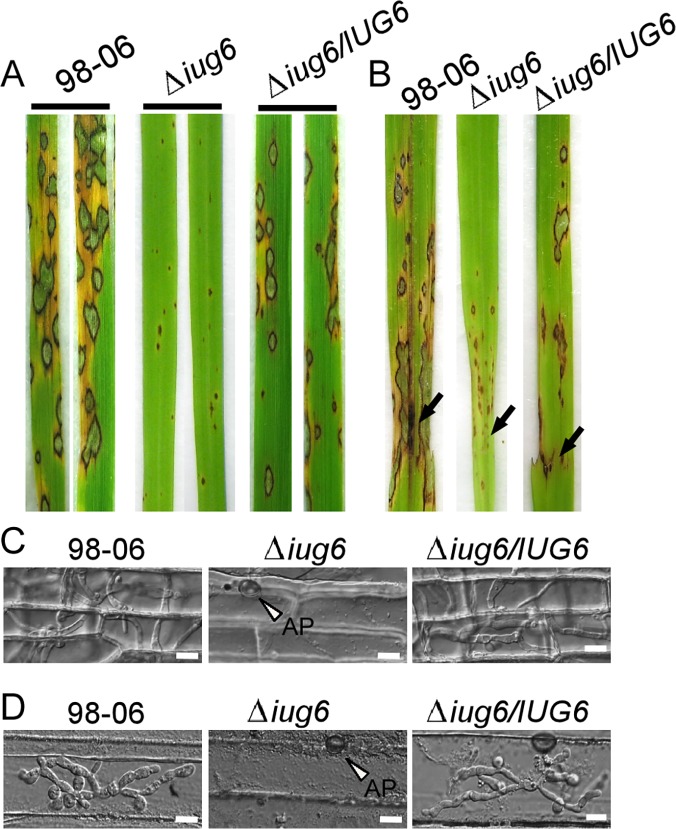
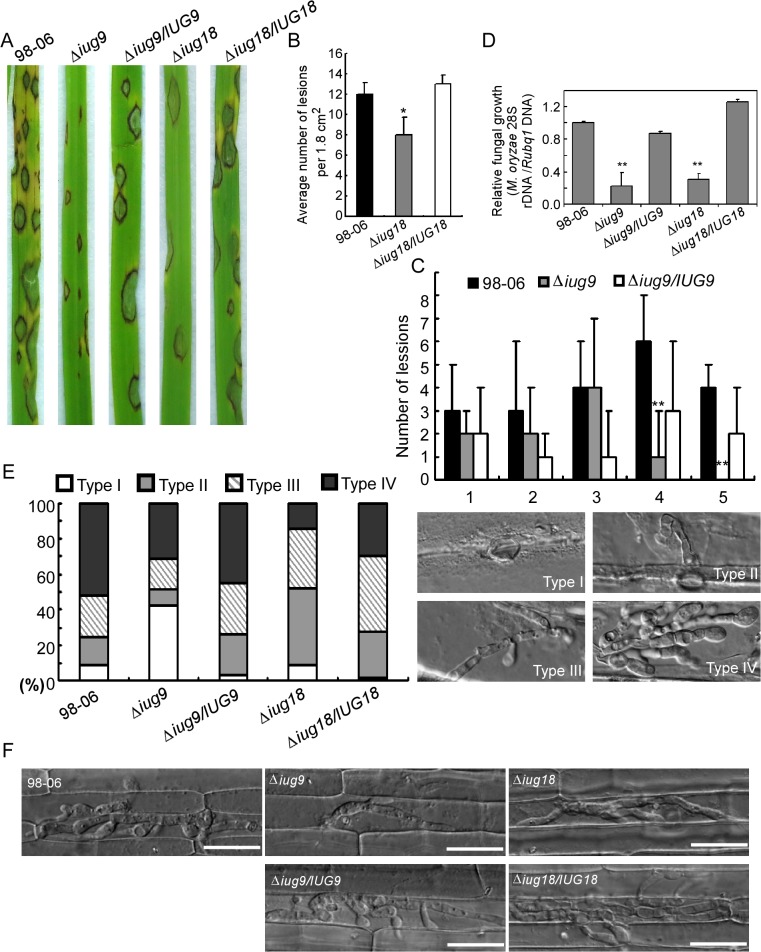
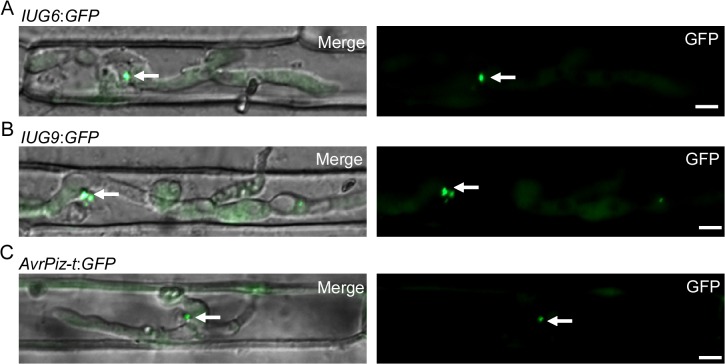
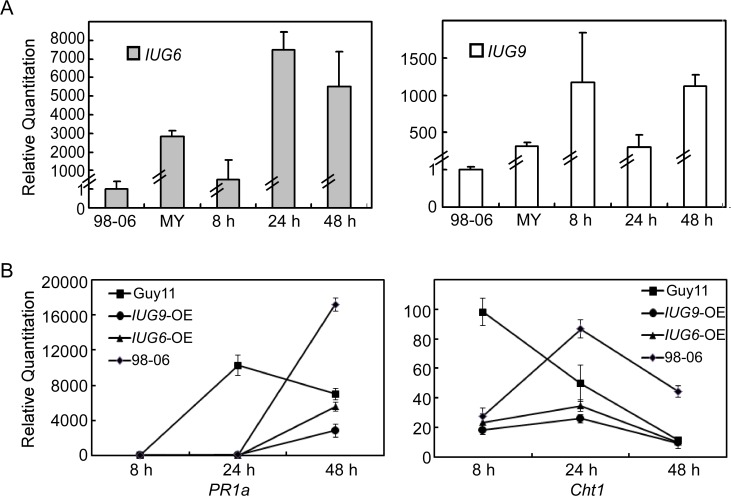
Genome dynamics of pathogenic organisms are driven by pathogen and host co-evolution, in which pathogen genomes are shaped to overcome stresses imposed by hosts with various genetic backgrounds through generation of a variety of isolates. This same principle applies to the rice blast pathogen Magnaporthe oryzae and the rice host; however, genetic variations among different isolates of M. oryzae remain largely unknown, particularly at genome and transcriptome levels. Here, we applied genomic and transcriptomic analytical tools to investigate M. oryzae isolate 98-06 that is the most aggressive in infection of susceptible rice cultivars. A unique 1.4 Mb of genomic sequences was found in isolate 98-06 in comparison to reference strain 70-15. Genome-wide expression profiling revealed the presence of two critical expression patterns of M. oryzae based on 64 known pathogenicity-related (PaR) genes. In addition, 134 candidate effectors with various segregation patterns were identified. Five tested proteins could suppress BAX-mediated programmed cell death in Nicotiana benthamiana leaves. Characterization of isolate-specific effector candidates Iug6 and Iug9 and PaR candidate Iug18 revealed that they have a role in fungal propagation and pathogenicity. Moreover, Iug6 and Iug9 are located exclusively in the biotrophic interfacial complex (BIC) and their overexpression leads to suppression of defense-related gene expression in rice, suggesting that they might participate in biotrophy by inhibiting the SA and ET pathways within the host. Thus, our studies identify novel effector and PaR proteins involved in pathogenicity of the highly aggressive M. oryzae field isolate 98-06, and reveal molecular and genomic dynamics in the evolution of M. oryzae and rice host interactions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Talbot NJ (2003) On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea . Annu Rev Microbiol 57: 177–202. - PubMed
-
- Ebbole DJ (2007) Magnaporthe as a model for understanding host-pathogen interactions. Annu Rev Phytopathol 45: 437–456. - PubMed
-
- Levy M, Correavictoria FJ, Zeigler RS, Xu SZ, Hamer JE (1993) Genetic Diversity of the Rice Blast Fungus in a Disease Nursery in Colombia. Phytopathology 83: 1427–1433.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
