

Epigenetics. Restricted epigenetic inheritance of H3K9 methylation
- PMID: 25838386
- PMCID: PMC4397586
- DOI: 10.1126/science.1260638
Epigenetics. Restricted epigenetic inheritance of H3K9 methylation
Abstract
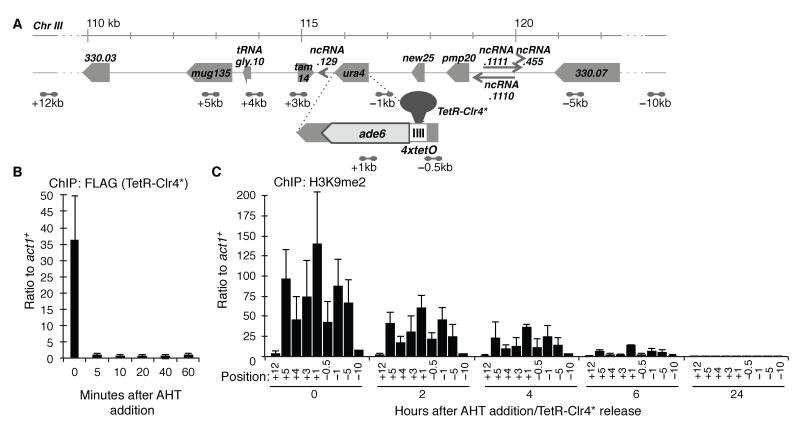
Posttranslational histone modifications are believed to allow the epigenetic transmission of distinct chromatin states, independently of associated DNA sequences. Histone H3 lysine 9 (H3K9) methylation is essential for heterochromatin formation; however, a demonstration of its epigenetic heritability is lacking. Fission yeast has a single H3K9 methyltransferase, Clr4, that directs all H3K9 methylation and heterochromatin. Using releasable tethered Clr4 reveals that an active process rapidly erases H3K9 methylation from tethering sites in wild-type cells. However, inactivation of the putative histone demethylase Epe1 allows H3K9 methylation and silent chromatin maintenance at the tethering site through many mitotic divisions, and transgenerationally through meiosis, after release of tethered Clr4. Thus, H3K9 methylation is a heritable epigenetic mark whose transmission is usually countered by its active removal, which prevents the unauthorized inheritance of heterochromatin.
Copyright © 2015, American Association for the Advancement of Science.
Figures



References
-
- Almouzni G, Probst AV. Heterochromatin maintenance and establishment: Lessons from the mouse pericentromere. nucleus. 2011;2:332–338. - PubMed
-
- Bannister AJ, et al. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
-
- Lachner M, O’Carroll D, Rea S, Mechtler K, Jenuwein T. Methylation of histone H3 lysine 9 creates a binding site for HP1 proteins. Nature. 2001;410:116–120. - PubMed
-
- Zhang K, Mosch K, Fischle W, Grewal S. Roles of the Clr4 methyltransferase complex in nucleation, spreading and maintenance of heterochromatin. Nat Struct Mol Biol. 2008;15:381–388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
