

Casein kinase 1δ is an APC/C(Cdh1) substrate that regulates cerebellar granule cell neurogenesis
- PMID: 25843713
- PMCID: PMC4401652
- DOI: 10.1016/j.celrep.2015.03.016
Casein kinase 1δ is an APC/C(Cdh1) substrate that regulates cerebellar granule cell neurogenesis
Abstract
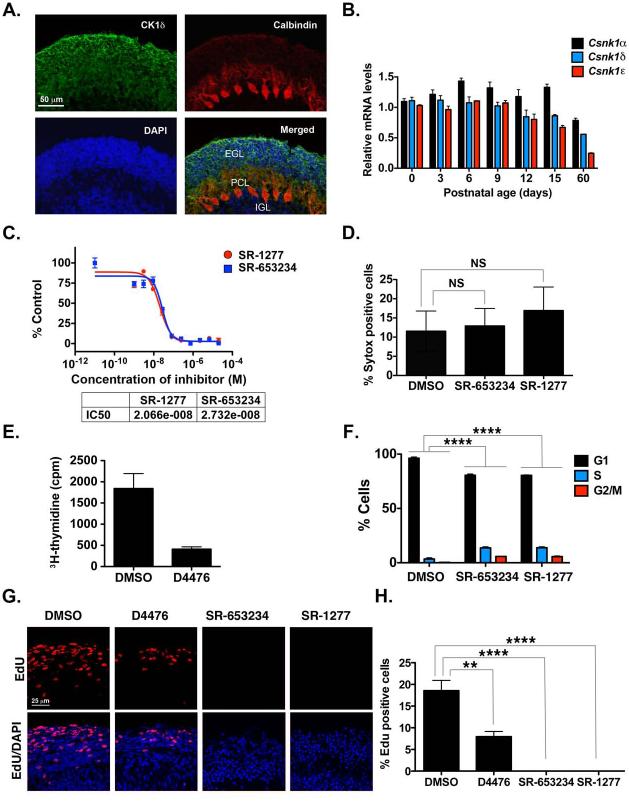
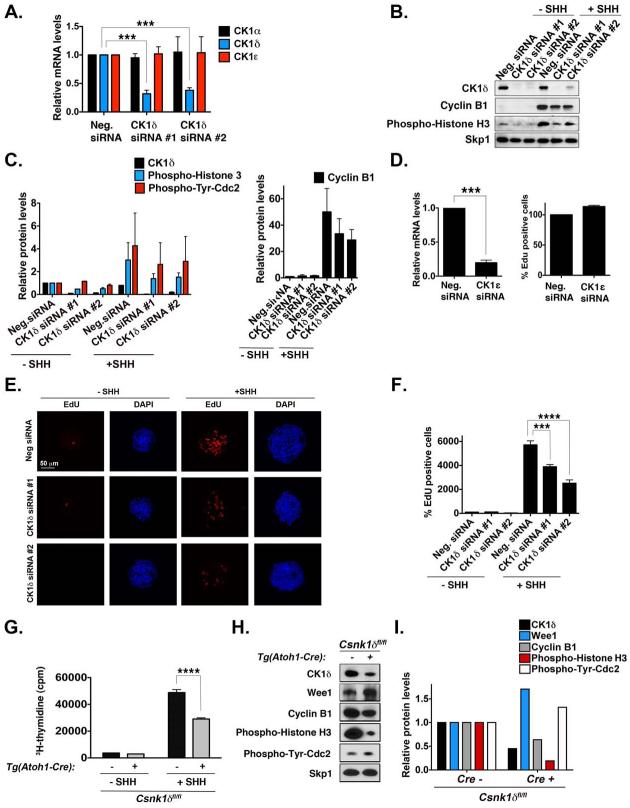
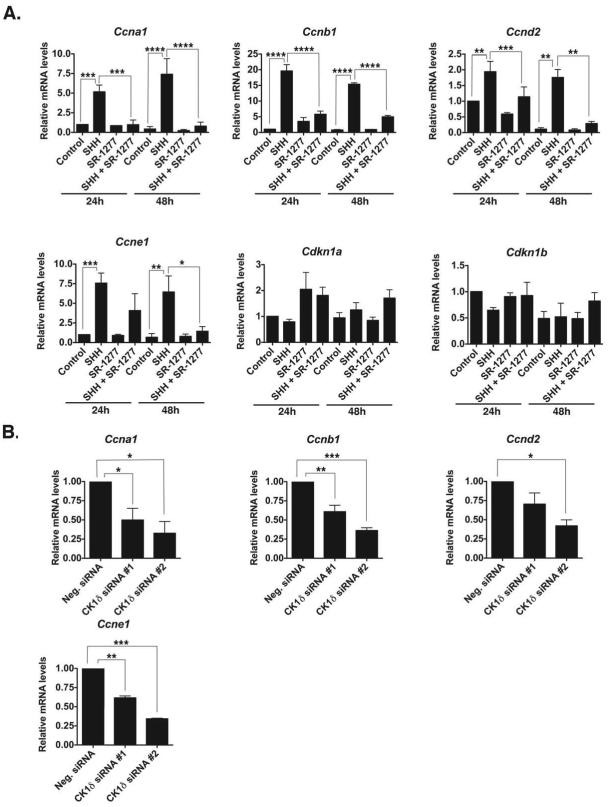
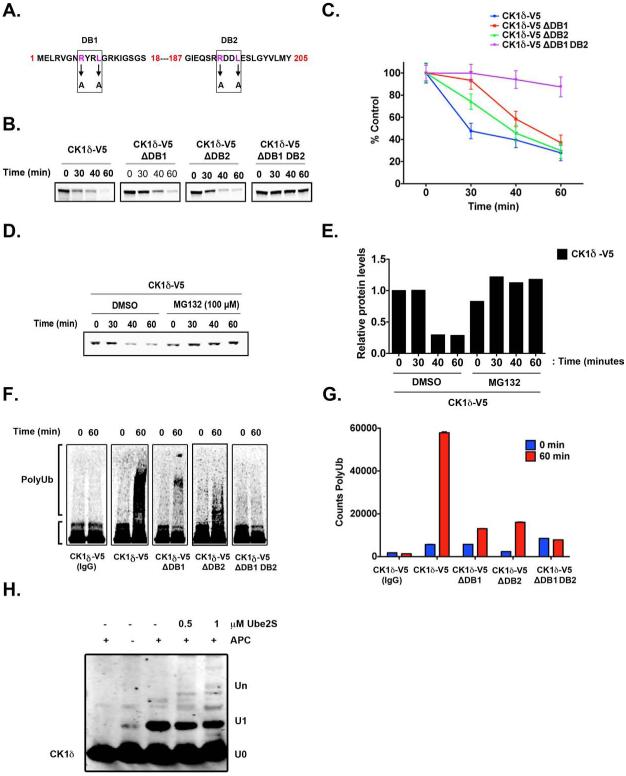
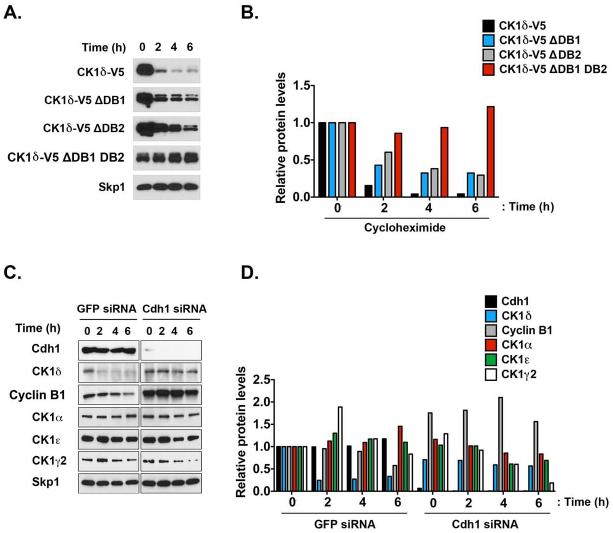
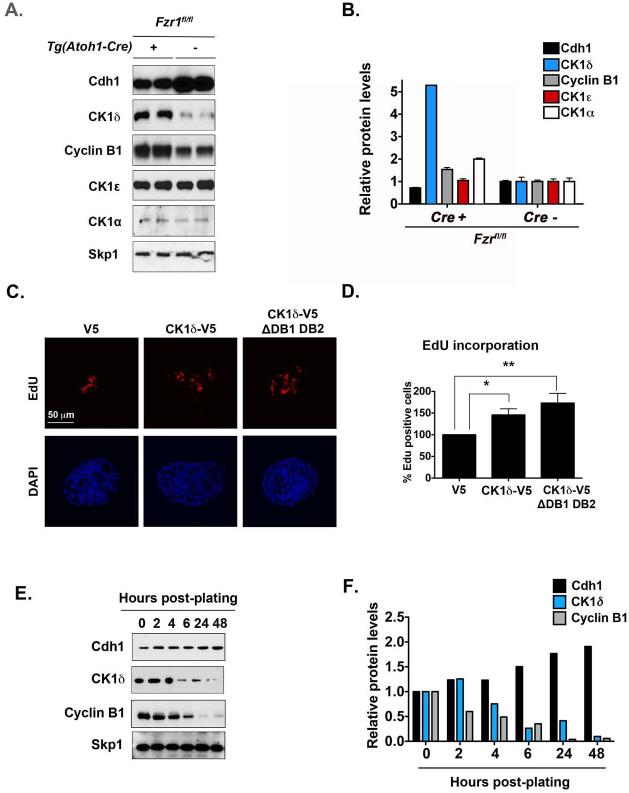
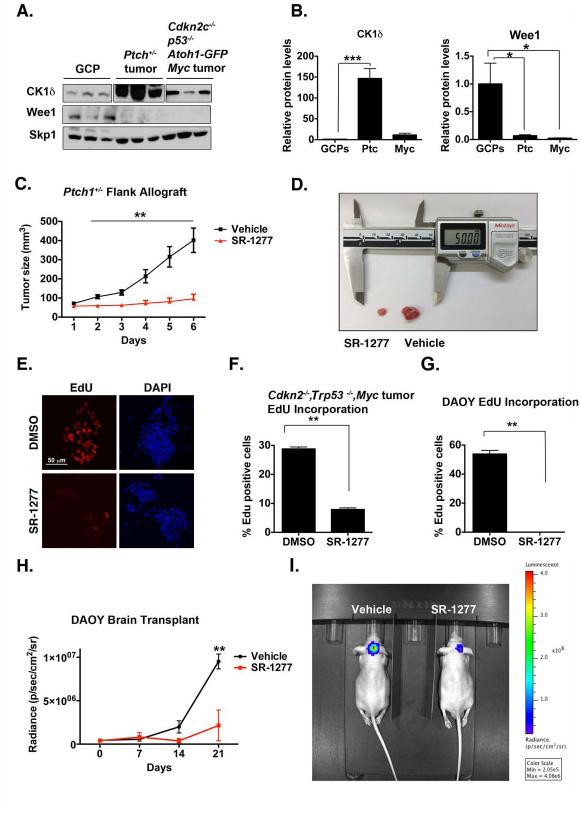
Although casein kinase 1δ (CK1δ) is at the center of multiple signaling pathways, its role in the expansion of CNS progenitor cells is unknown. Using mouse cerebellar granule cell progenitors (GCPs) as a model for brain neurogenesis, we demonstrate that the loss of CK1δ or treatment of GCPs with a highly selective small molecule inhibits GCP expansion. In contrast, CK1δ overexpression increases GCP proliferation. Thus, CK1δ appears to regulate GCP neurogenesis. CK1δ is targeted for proteolysis via the anaphase-promoting complex/cyclosome (APC/C(Cdh1)) ubiquitin ligase, and conditional deletion of the APC/C(Cdh1) activator Cdh1 in cerebellar GCPs results in higher levels of CK1δ. APC/C(Cdh1) also downregulates CK1δ during cell-cycle exit. Therefore, we conclude that APC/C(Cdh1) controls CK1δ levels to balance proliferation and cell-cycle exit in the developing CNS. Similar studies in medulloblastoma cells showed that CK1δ holds promise as a therapeutic target.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Barford D. Structure, function and mechanism of the anaphase promoting complex (APC/C) Q. Rev. Biophys. 2011;44:153–190. - PubMed
-
- Behrend L, Stoter M, Kurth M, Rutter G, Heukeshoven J, Deppert W, Knippschild U. Interaction of casein kinase 1 delta (CK1delta) with post-Golgi structures, microtubules and the spindle apparatus. Eur. J. C.ell Biol. 2000;79:240–251. - PubMed
-
- Ben-Arie N, Bellen HJ, Armstrong DL, McCall AE, Gordadze PR, Guo Q, Matzuk MM, Zoghbi HY. Math1 is essential for genesis of cerebellar granule neurons. Nature. 1997;390:169–172. - PubMed
-
- Beyaert R, Vanhaesebroeck B, Declercq W, Van Lint J, Vandenabele P, Agostinis P, Vandenheede JR, Fiers W. Casein kinase-1 phosphorylates the p75 tumor necrosis factor receptor and negatively regulates tumor necrosis factor signaling for apoptosis. J. Biol. Chem. 1995;270:23293–23299. - PubMed
-
- Bibian M, Rahaim RJ, Choi JY, Noguchi Y, Schurer S, Chen W, Nakanishi S, Licht K, Rosenberg LH, Li L, et al. Development of highly selective casein kinase 1delta/1epsilon (CK1delta/epsilon) inhibitors with potent antiproliferative properties. Bioorg. Med. Chem. Lett. 2013;23:4374–4380. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
