

Molecular crowding enhances facilitated diffusion of two human DNA glycosylases
- PMID: 25845592
- PMCID: PMC4417188
- DOI: 10.1093/nar/gkv301
Molecular crowding enhances facilitated diffusion of two human DNA glycosylases
Abstract
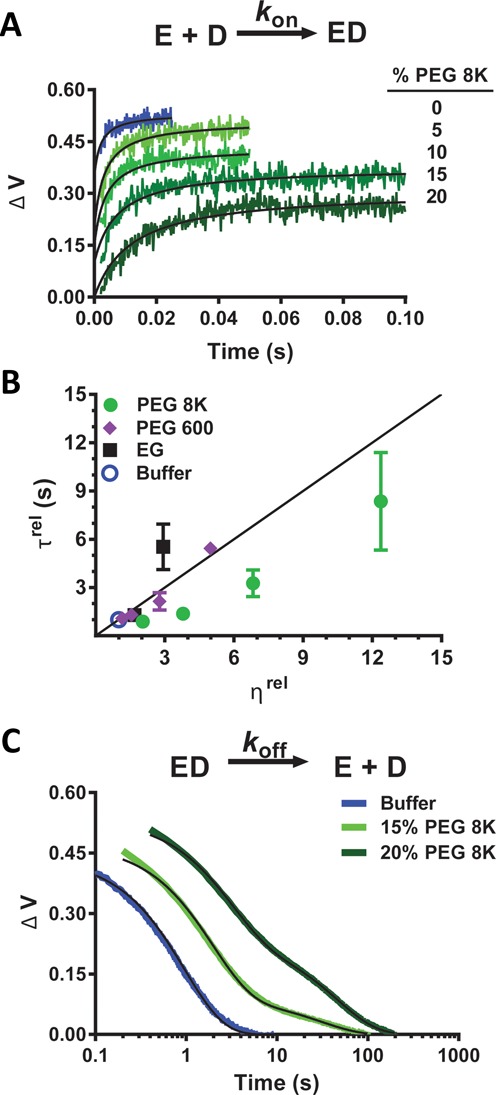
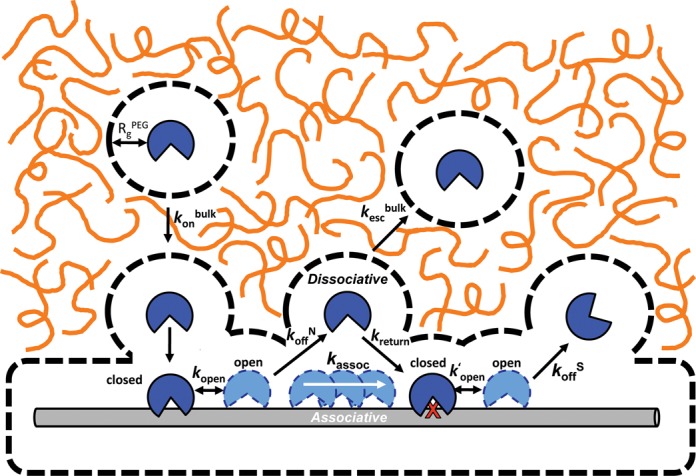
Intracellular space is at a premium due to the high concentrations of biomolecules and is expected to have a fundamental effect on how large macromolecules move in the cell. Here, we report that crowded solutions promote intramolecular DNA translocation by two human DNA repair glycosylases. The crowding effect increases both the efficiency and average distance of DNA chain translocation by hindering escape of the enzymes to bulk solution. The increased contact time with the DNA chain provides for redundant damage patrolling within individual DNA chains at the expense of slowing the overall rate of damaged base removal from a population of molecules. The significant biological implication is that a crowded cellular environment could influence the mechanism of damage recognition as much as any property of the enzyme or DNA.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
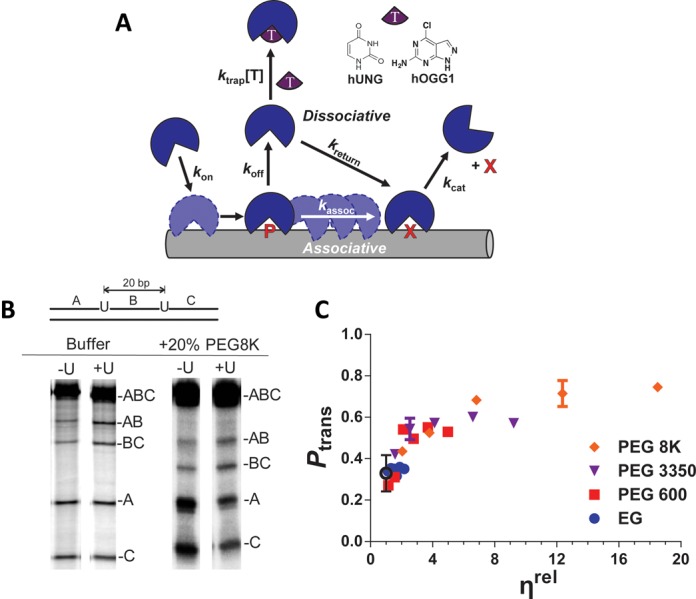
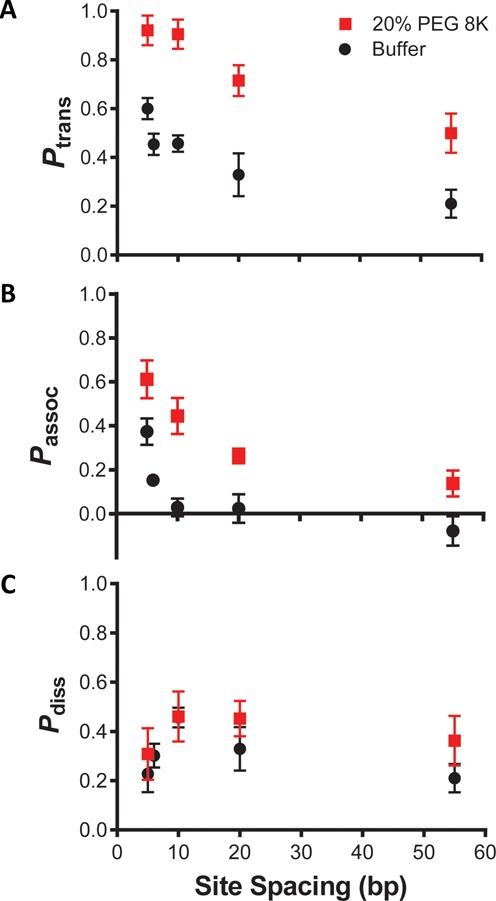
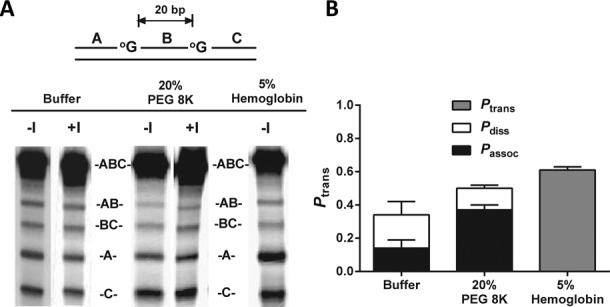
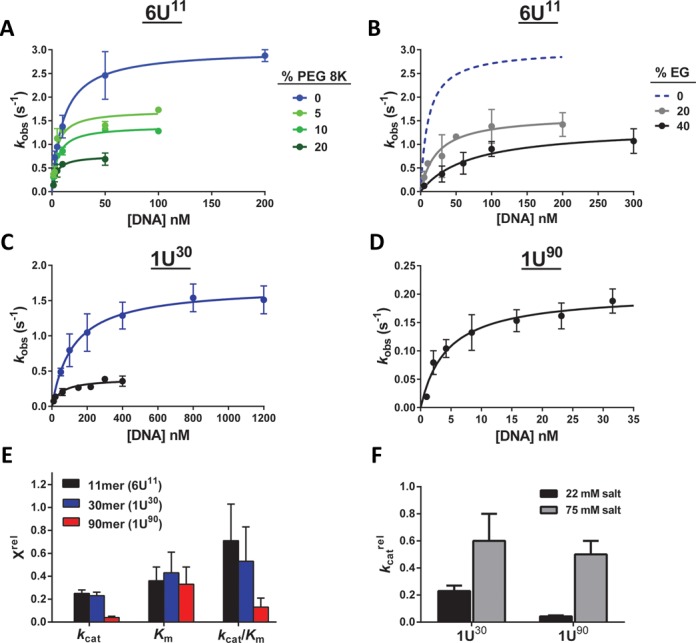
References
-
- Krosky D.J., Song F., Stivers J.T. The origins of high-affinity enzyme binding to an extrahelical DNA base. Biochemistry. 2005;44:5949–5959. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
