

Extracellular zinc induces phosphoethanolamine addition to Pseudomonas aeruginosa lipid A via the ColRS two-component system
- PMID: 25846400
- PMCID: PMC4715879
- DOI: 10.1111/mmi.13018
Extracellular zinc induces phosphoethanolamine addition to Pseudomonas aeruginosa lipid A via the ColRS two-component system
Abstract
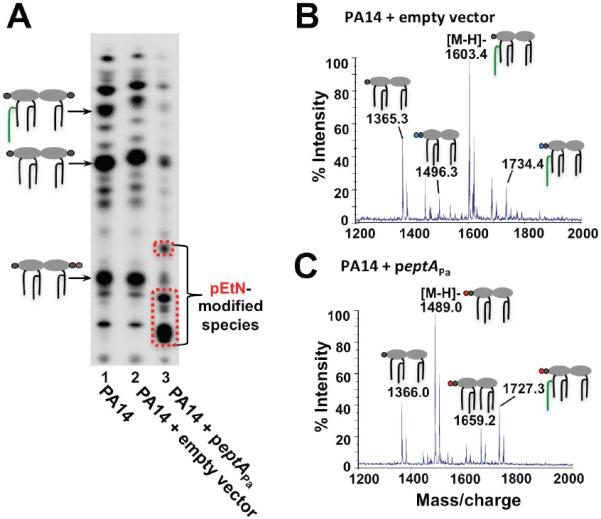
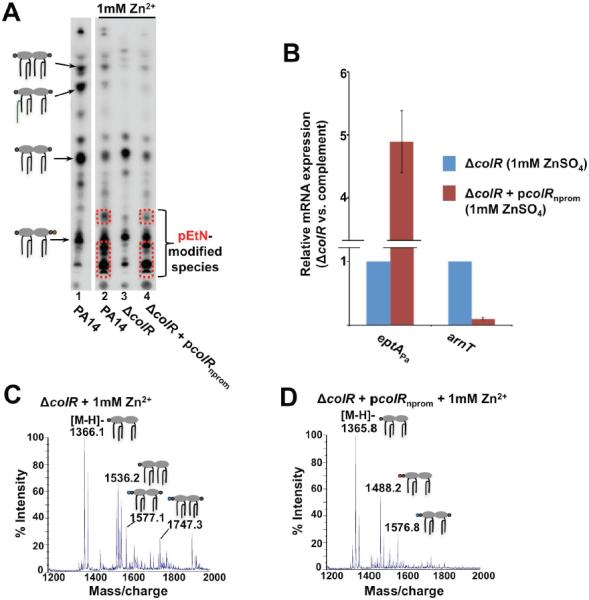
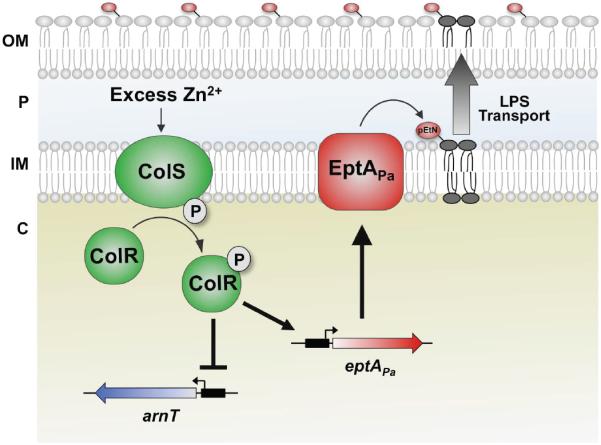
Gram-negative bacteria survive harmful environmental stressors by modifying their outer membrane. Much of this protection is afforded upon remodeling of the lipid A region of the major surface molecule lipopolysaccharide (LPS). For example, the addition of cationic substituents, such as 4-amino-4-deoxy-L-arabinose (L-Ara4N) and phosphoehthanolamine (pEtN) at the lipid A phosphate groups, is often induced in response to specific environmental flux stabilizing the outer membrane. The work herein represents the first report of pEtN addition to Pseudomonas aeruginosa lipid A. We have identified the key pEtN transferase which we named EptAPa and characterized its strict activity on only one position of lipid A, contrasting from previously studied EptA enzymes. We further show that transcription of eptAP a is regulated by zinc via the ColRS two-component system instead of the PmrAB system responsible for eptA regulation in E. coli and Salmonella enterica. Further, although L-Ara4N is readily added to the same position of lipid A as pEtN under certain environmental conditions, ColR specifically induces pEtN addition to lipid A in lieu of L-Ara4N when Zn2+ is present. The unique, specific regulation of eptAP a transcription and enzymatic activity described in this work demonstrates the tight yet inducible control over LPS modification in P. aeruginosa.
© 2015 John Wiley & Sons Ltd.
Figures




References
-
- Andreini C, Bertini I, Cavallaro G, Holliday GL, Thornton JM. Metal ions in biological catalysis: from enzyme databases to general principles. JBIC J Biol Inorg Chem. 2008;13:1205–1218. - PubMed
-
- Bader MW, Sanowar S, Daley ME, Schneider AR, Cho U, Xu W, et al. Recognition of Antimicrobial Peptides by a Bacterial Sensor Kinase. Cell. 2005;122:461–472. - PubMed
-
- Ballesta S, Conejo MC, García I, Rodríguez-Martínez JM, Velasco C, Pascual A. Survival and resistance to imipenem of Pseudomonas aeruginosa on latex gloves. J Antimicrob Chemother. 2006;57:1010–1012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
