

Maintenance of distinct melanocyte populations in the interfollicular epidermis
- PMID: 25847135
- PMCID: PMC4973853
- DOI: 10.1111/pcmr.12375
Maintenance of distinct melanocyte populations in the interfollicular epidermis
Abstract
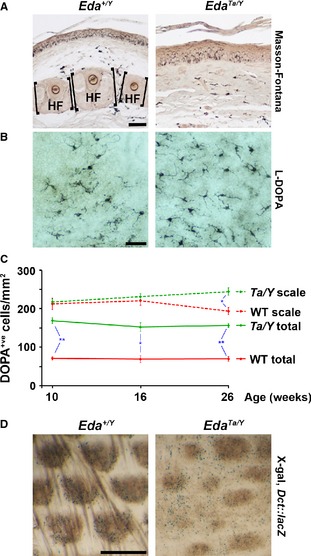
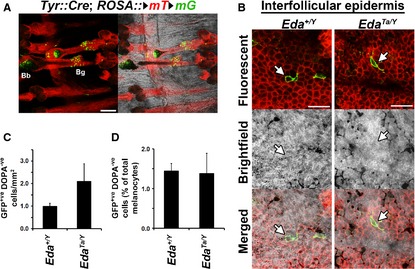
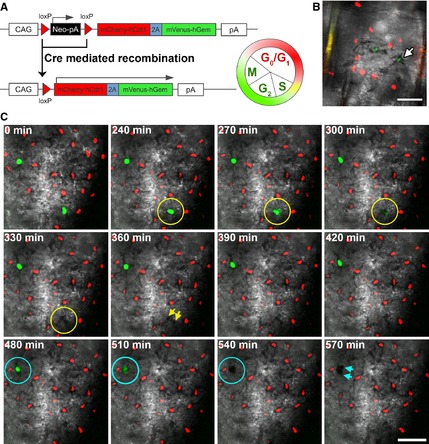
Hair follicles and sweat glands are recognized as reservoirs of melanocyte stem cells (MSCs). Unlike differentiated melanocytes, undifferentiated MSCs do not produce melanin. They serve as a source of differentiated melanocytes for the hair follicle and contribute to the interfollicular epidermis upon wounding, exposure to ultraviolet irradiation or in remission from vitiligo, where repigmentation often spreads outwards from the hair follicles. It is unknown whether these observations reflect the normal homoeostatic mechanism of melanocyte renewal or whether unperturbed interfollicular epidermis can maintain a melanocyte population that is independent of the skin's appendages. Here, we show that mouse tail skin lacking appendages does maintain a stable melanocyte number, including a low frequency of amelanotic melanocytes, into adult life. Furthermore, we show that actively cycling differentiated melanocytes are present in postnatal skin, indicating that amelanotic melanocytes are not uniquely relied on for melanocyte homoeostasis.
Keywords: epidermis; hair follicle; interfollicular; melanocyte; stem cell.
© 2015 The Authors. Pigment Cell & Melanoma Research Published by John Wiley & Sons Ltd.
Figures
Similar articles
-
Different populations of melanocytes are present in hair follicles and epidermis.Pigment Cell Res. 1996 Dec;9(6):304-10. doi: 10.1111/j.1600-0749.1996.tb00122.x. Pigment Cell Res. 1996. PMID: 9125754
-
Isolation of Epidermal and Hair Follicle Melanocytes.Methods Mol Biol. 2020;2154:23-32. doi: 10.1007/978-1-0716-0648-3_3. Methods Mol Biol. 2020. PMID: 32314205
-
Melanocyte stem cells: a melanocyte reservoir in hair follicles for hair and skin pigmentation.Pigment Cell Melanoma Res. 2011 Jun;24(3):401-10. doi: 10.1111/j.1755-148X.2011.00855.x. Epub 2011 May 5. Pigment Cell Melanoma Res. 2011. PMID: 21466661 Review.
-
Repigmentation through Melanocyte Regeneration in Vitiligo.Dermatol Clin. 2017 Apr;35(2):205-218. doi: 10.1016/j.det.2016.11.015. Dermatol Clin. 2017. PMID: 28317529 Review.
-
Repigmentation of Human Vitiligo Skin by NBUVB Is Controlled by Transcription of GLI1 and Activation of the β-Catenin Pathway in the Hair Follicle Bulge Stem Cells.J Invest Dermatol. 2018 Mar;138(3):657-668. doi: 10.1016/j.jid.2017.09.040. Epub 2017 Oct 17. J Invest Dermatol. 2018. PMID: 29054607 Free PMC article.
Cited by
-
Spatiotemporal Labeling of Melanocytes in Mice.Int J Mol Sci. 2018 May 15;19(5):1469. doi: 10.3390/ijms19051469. Int J Mol Sci. 2018. PMID: 29762513 Free PMC article. Review.
-
The journey from melanocytes to melanoma.Nat Rev Cancer. 2023 Jun;23(6):372-390. doi: 10.1038/s41568-023-00565-7. Epub 2023 Apr 24. Nat Rev Cancer. 2023. PMID: 37095242 Review.
-
Dermal papilla cells and melanocytes response to physiological oxygen levels depends on their interactions.Cell Prolif. 2021 Jul;54(7):e13013. doi: 10.1111/cpr.13013. Epub 2021 Jun 8. Cell Prolif. 2021. PMID: 34101928 Free PMC article.
-
The convergence theory for vitiligo: A reappraisal.Exp Dermatol. 2019 Jun;28(6):647-655. doi: 10.1111/exd.13677. Epub 2018 Jun 28. Exp Dermatol. 2019. PMID: 29704874 Free PMC article. Review.
-
Understanding Mammalian Hair Follicle Ecosystems by Single-Cell RNA Sequencing.Animals (Basel). 2022 Sep 14;12(18):2409. doi: 10.3390/ani12182409. Animals (Basel). 2022. PMID: 36139270 Free PMC article. Review.
References
-
- Delmas, V. , Martinozzi, S. , Bourgeois, Y. , Holzenberger, M. , and Larue, L. (2003). Cre‐mediated recombination in the skin melanocyte lineage. Genesis 36, 73–80. - PubMed
-
- Falconer, D.S. (1953). Total sex‐linkage in the house mouse. Z. Indukt. Abstamm.‐u. Vererb.‐L. 85, 210–219. - PubMed
-
- Grichnik, J.M. , Ali, W.N. , Burch, J.A. , Byers, J.D. , Garcia, C.A. , Clark, R.E. , and Shea, C.R. (1996). KIT expression reveals a population of precursor melanocytes in human skin. J. Invest. Dermatol. 106, 967–971. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
