

Diaminopimelic Acid Amidation in Corynebacteriales: NEW INSIGHTS INTO THE ROLE OF LtsA IN PEPTIDOGLYCAN MODIFICATION
- PMID: 25847251
- PMCID: PMC4505564
- DOI: 10.1074/jbc.M115.642843
Diaminopimelic Acid Amidation in Corynebacteriales: NEW INSIGHTS INTO THE ROLE OF LtsA IN PEPTIDOGLYCAN MODIFICATION
Abstract
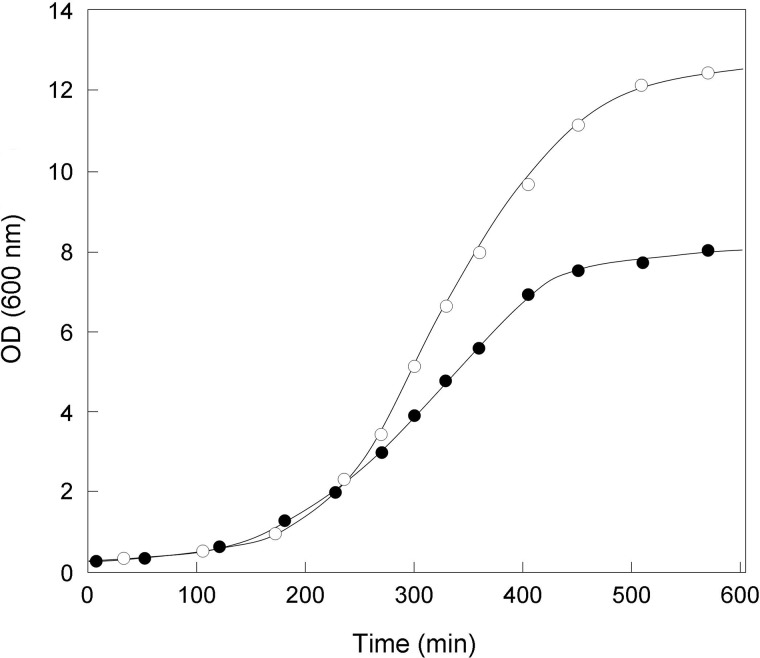
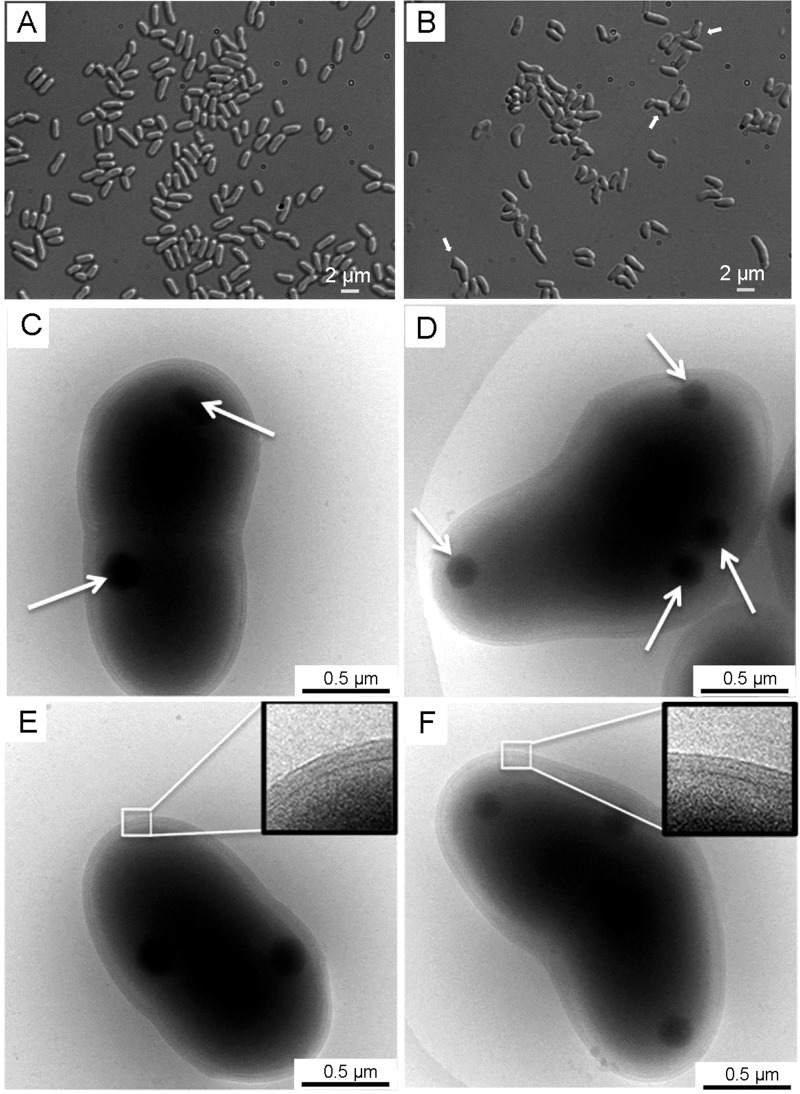
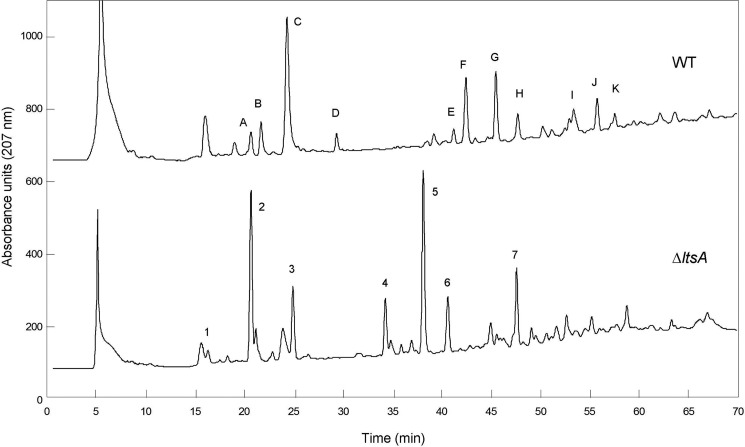
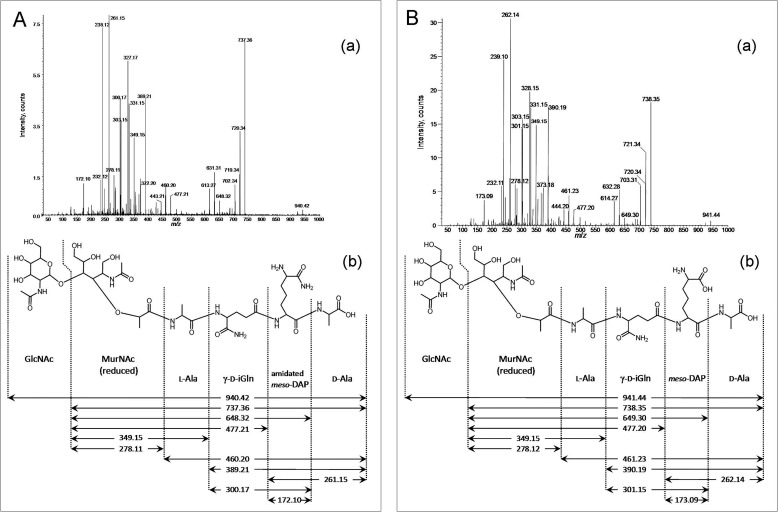
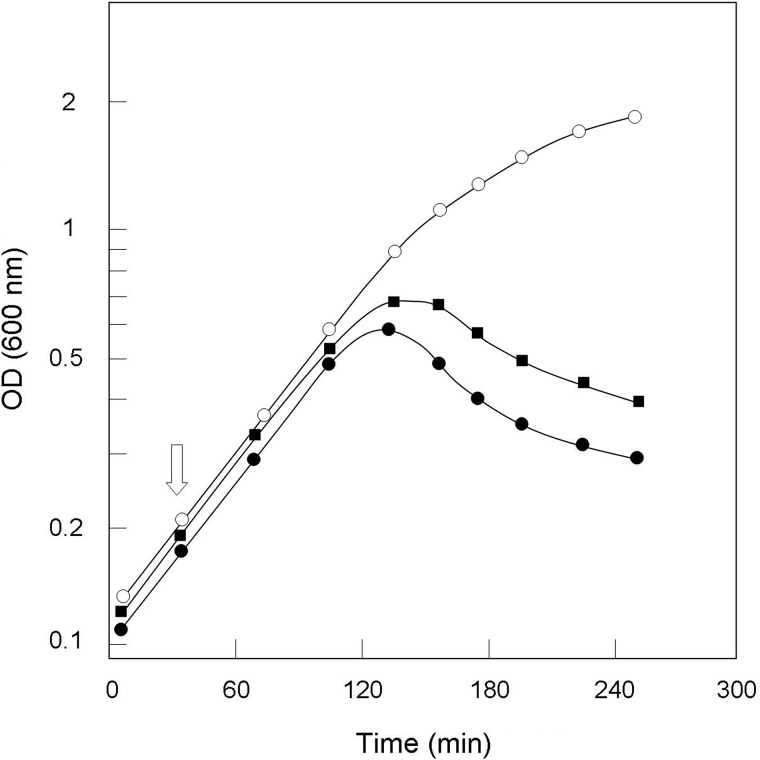
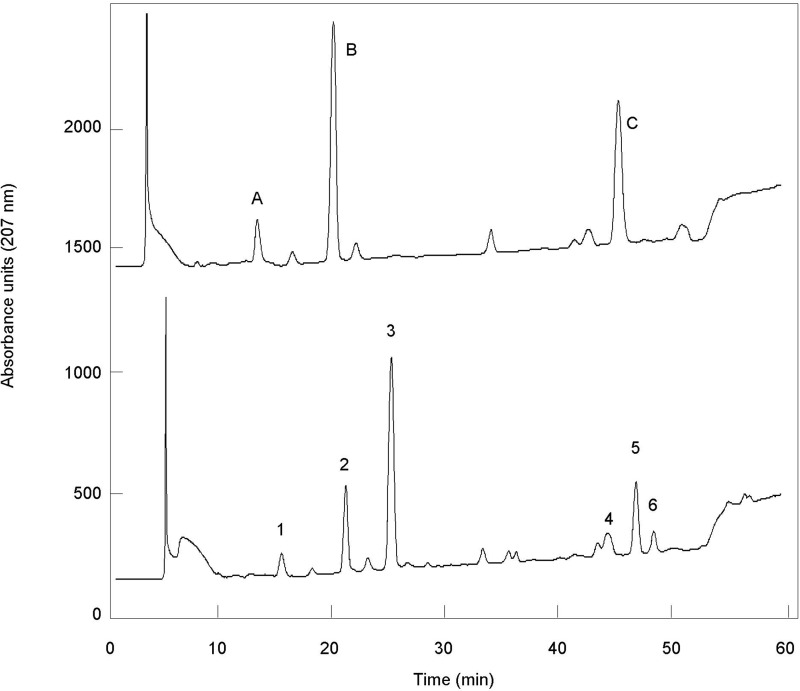
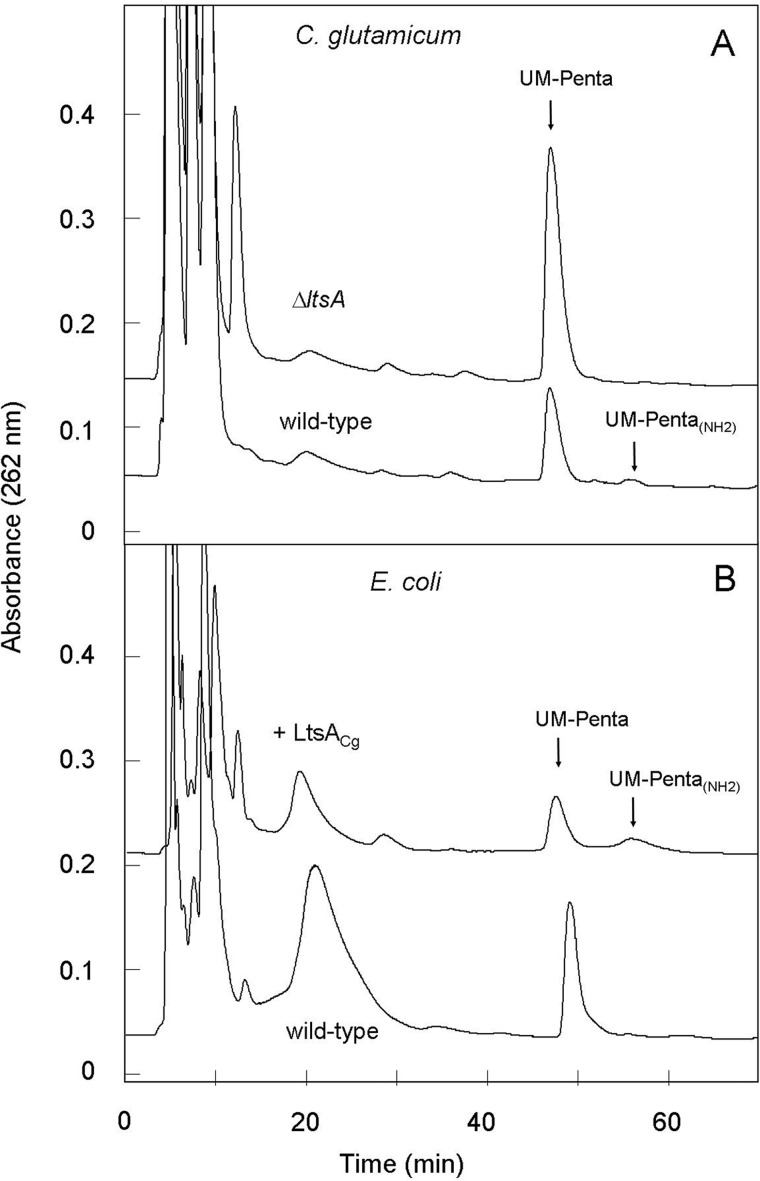
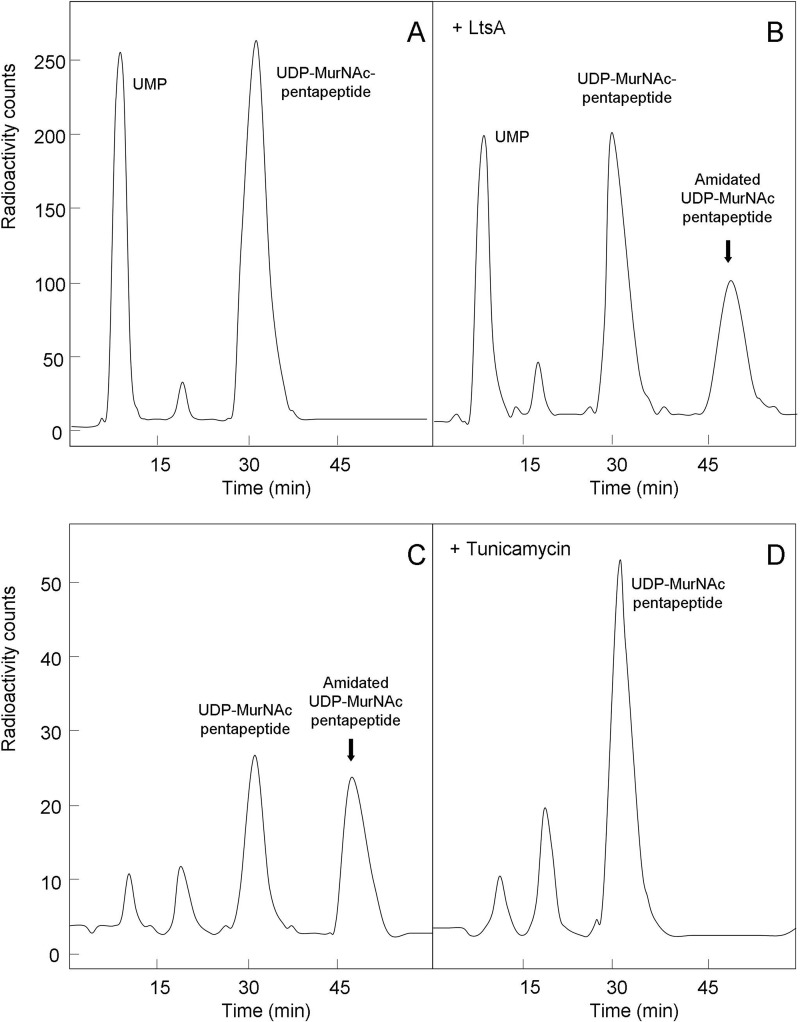
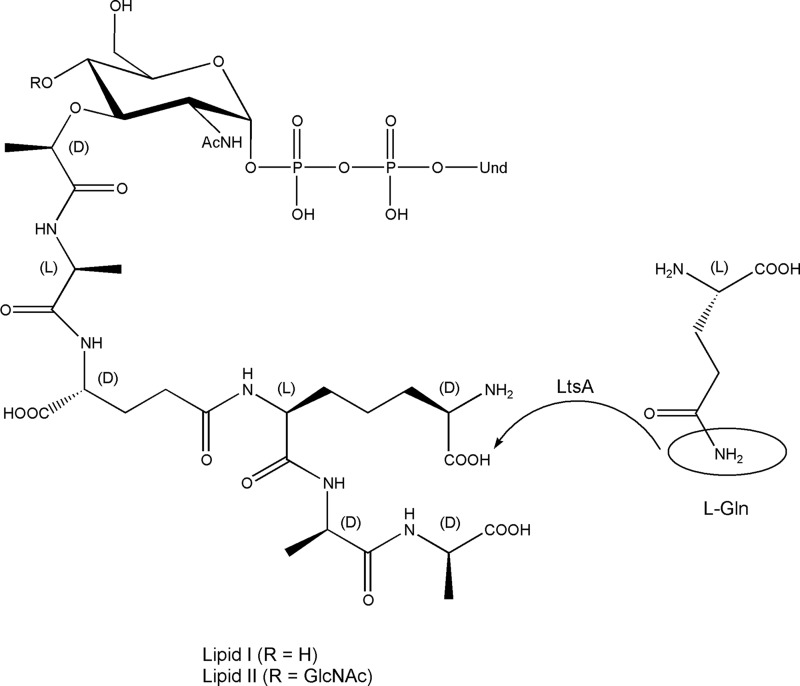
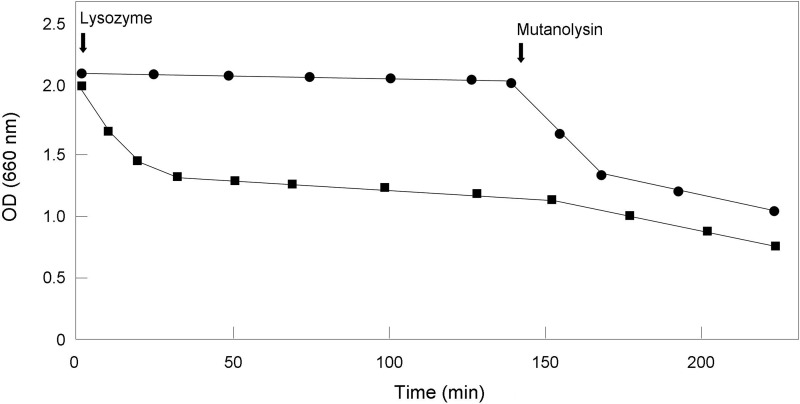
A gene named ltsA was earlier identified in Rhodococcus and Corynebacterium species while screening for mutations leading to increased cell susceptibility to lysozyme. The encoded protein belonged to a huge family of glutamine amidotransferases whose members catalyze amide nitrogen transfer from glutamine to various specific acceptor substrates. We here describe detailed physiological and biochemical investigations demonstrating the specific role of LtsA protein from Corynebacterium glutamicum (LtsACg) in the modification by amidation of cell wall peptidoglycan diaminopimelic acid (DAP) residues. A morphologically altered but viable ΔltsA mutant was generated, which displays a high susceptibility to lysozyme and β-lactam antibiotics. Analysis of its peptidoglycan structure revealed a total loss of DAP amidation, a modification that was found in 80% of DAP residues in the wild-type polymer. The cell peptidoglycan content and cross-linking were otherwise not modified in the mutant. Heterologous expression of LtsACg in Escherichia coli yielded a massive and toxic incorporation of amidated DAP into the peptidoglycan that ultimately led to cell lysis. In vitro assays confirmed the amidotransferase activity of LtsACg and showed that this enzyme used the peptidoglycan lipid intermediates I and II but not, or only marginally, the UDP-MurNAc pentapeptide nucleotide precursor as acceptor substrates. As is generally the case for glutamine amidotransferases, either glutamine or NH4(+) could serve as the donor substrate for LtsACg. The enzyme did not amidate tripeptide- and tetrapeptide-truncated versions of lipid I, indicating a strict specificity for a pentapeptide chain length.
Keywords: Corynebacteriales; DAP amidation; antibiotics; bacterial metabolism; cell wall; enzyme; gene knockout; glutaminase; lysozyme; peptidoglycan.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

Similar articles
-
Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum.Microorganisms. 2021 Feb 16;9(2):409. doi: 10.3390/microorganisms9020409. Microorganisms. 2021. PMID: 33669405 Free PMC article.
-
Different modes of diaminopimelate synthesis and their role in cell wall integrity: a study with Corynebacterium glutamicum.J Bacteriol. 1998 Jun;180(12):3159-65. doi: 10.1128/JB.180.12.3159-3165.1998. J Bacteriol. 1998. PMID: 9620966 Free PMC article.
-
A mutation in the Corynebacterium glutamicum ltsA gene causes susceptibility to lysozyme, temperature-sensitive growth, and L-glutamate production.J Bacteriol. 2000 May;182(10):2696-701. doi: 10.1128/JB.182.10.2696-2701.2000. J Bacteriol. 2000. PMID: 10781535 Free PMC article.
-
Lysozyme substrates.EXS. 1996;75:105-10. doi: 10.1007/978-3-0348-9225-4_7. EXS. 1996. PMID: 8765297 Review.
-
Metabolic labeling probes for interrogation of the host-pathogen interaction.Org Biomol Chem. 2021 Apr 7;19(13):2856-2870. doi: 10.1039/d0ob02517h. Epub 2021 Mar 16. Org Biomol Chem. 2021. PMID: 33725048 Review.
Cited by
-
The impact of co-fed plastic diet on Tenebrio molitor gut bacterial community structure.Sci Rep. 2025 Jul 1;15(1):21203. doi: 10.1038/s41598-025-04805-8. Sci Rep. 2025. PMID: 40594024 Free PMC article.
-
Amidation of glutamate residues in mycobacterial peptidoglycan is essential for cell wall cross-linking.Front Cell Infect Microbiol. 2023 Aug 24;13:1205829. doi: 10.3389/fcimb.2023.1205829. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37692163 Free PMC article.
-
Facile Synthesis and Metabolic Incorporation of m-DAP Bioisosteres Into Cell Walls of Live Bacteria.ACS Chem Biol. 2020 Nov 20;15(11):2966-2975. doi: 10.1021/acschembio.0c00618. Epub 2020 Oct 20. ACS Chem Biol. 2020. PMID: 33078931 Free PMC article.
-
Proteome Changes Induced by Iprodione Exposure in the Pesticide-Tolerant Pseudomonas sp. C9 Strain Isolated from a Biopurification System.Int J Mol Sci. 2024 Sep 28;25(19):10471. doi: 10.3390/ijms251910471. Int J Mol Sci. 2024. PMID: 39408799 Free PMC article.
-
Requirement of the LtsA Protein for Formation of the Mycolic Acid-Containing Layer on the Cell Surface of Corynebacterium glutamicum.Microorganisms. 2021 Feb 16;9(2):409. doi: 10.3390/microorganisms9020409. Microorganisms. 2021. PMID: 33669405 Free PMC article.
References
-
- Crick D. C., Mahapatra S., Brennan P. J. (2001) Biosynthesis of the arabinogalactan-peptidoglycan complex of Mycobacterium tuberculosis. Glycobiology 11, 107R–118R - PubMed
-
- Puech V., Chami M., Lemassu A., Lanéelle M. A., Schiffler B., Gounon P., Bayan N., Benz R., Daffé M. (2001) Structure of the cell envelope of corynebacteria: importance of the non-covalently bound lipids in the formation of the cell wall permeability barrier and fracture plane. Microbiology 147, 1365–1382 - PubMed
-
- McNeil M., Daffé M., Brennan P. J. (1990) Evidence for the nature of the link between the arabinogalactan and peptidoglycan of mycobacterial cell walls. J. Biol. Chem. 265, 18200–18206 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
