

Etiology and pathogenesis of the cohesinopathies
- PMID: 25847322
- PMCID: PMC6680315
- DOI: 10.1002/wdev.190
Etiology and pathogenesis of the cohesinopathies
Abstract
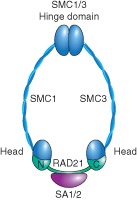
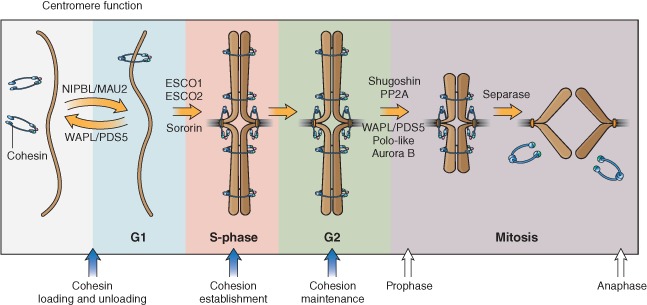
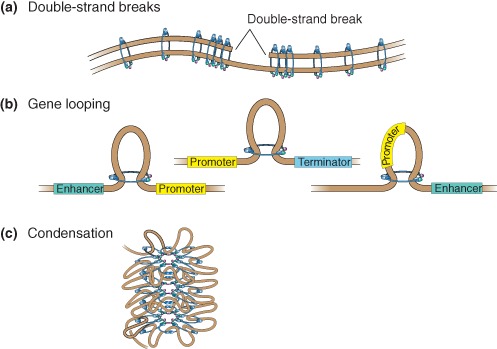
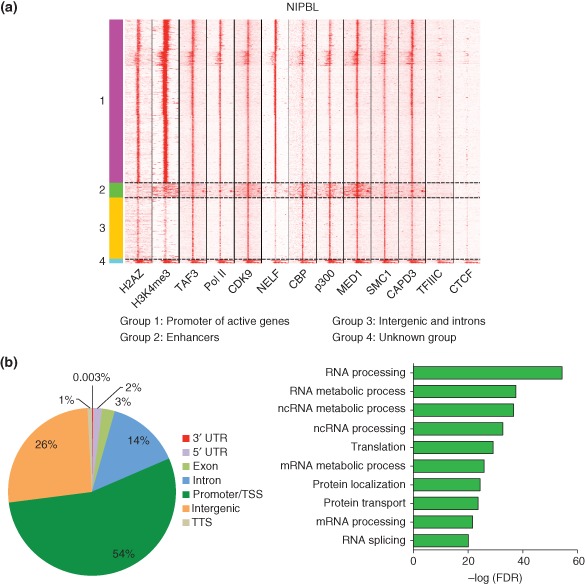
Cohesin is a chromosome-associated protein complex that plays many important roles in chromosome function. Genetic screens in yeast originally identified cohesin as a key regulator of chromosome segregation. Subsequently, work by various groups has identified cohesin as critical for additional processes such as DNA damage repair, insulator function, gene regulation, and chromosome condensation. Mutations in the genes encoding cohesin and its accessory factors result in a group of developmental and intellectual impairment diseases termed 'cohesinopathies.' How mutations in cohesin genes cause disease is not well understood as precocious chromosome segregation is not a common feature in cells derived from patients with these syndromes. In this review, the latest findings concerning cohesin's function in the organization of chromosome structure and gene regulation are discussed. We propose that the cohesinopathies are caused by changes in gene expression that can negatively impact translation. The similarities and differences between cohesinopathies and ribosomopathies, diseases caused by defects in ribosome biogenesis, are discussed. The contribution of cohesin and its accessory proteins to gene expression programs that support translation suggests that cohesin provides a means of coupling chromosome structure with the translational output of cells.
© 2015 The Authors. WIREs Developmental Biology published by Wiley Periodicals, Inc.
Figures
Similar articles
-
Translational mechanisms at work in the cohesinopathies.Nucleus. 2012 Nov-Dec;3(6):520-5. doi: 10.4161/nucl.22800. Epub 2012 Nov 1. Nucleus. 2012. PMID: 23138777 Free PMC article.
-
Cohesin proteins promote ribosomal RNA production and protein translation in yeast and human cells.PLoS Genet. 2012;8(6):e1002749. doi: 10.1371/journal.pgen.1002749. Epub 2012 Jun 14. PLoS Genet. 2012. PMID: 22719263 Free PMC article.
-
The roles of cohesins in mitosis, meiosis, and human health and disease.Methods Mol Biol. 2014;1170:229-66. doi: 10.1007/978-1-4939-0888-2_11. Methods Mol Biol. 2014. PMID: 24906316 Free PMC article. Review.
-
Stimulation of mTORC1 with L-leucine rescues defects associated with Roberts syndrome.PLoS Genet. 2013;9(10):e1003857. doi: 10.1371/journal.pgen.1003857. Epub 2013 Oct 3. PLoS Genet. 2013. PMID: 24098154 Free PMC article.
-
Cohesinopathies of a feather flock together.PLoS Genet. 2013;9(12):e1004036. doi: 10.1371/journal.pgen.1004036. Epub 2013 Dec 19. PLoS Genet. 2013. PMID: 24367282 Free PMC article. Review.
Cited by
-
Role of the DDX11 DNA Helicase in Warsaw Breakage Syndrome Etiology.Int J Mol Sci. 2021 Feb 25;22(5):2308. doi: 10.3390/ijms22052308. Int J Mol Sci. 2021. PMID: 33669056 Free PMC article. Review.
-
The Interplay of Cohesin and RNA Processing Factors: The Impact of Their Alterations on Genome Stability.Int J Mol Sci. 2022 Apr 1;23(7):3939. doi: 10.3390/ijms23073939. Int J Mol Sci. 2022. PMID: 35409298 Free PMC article. Review.
-
Non-redundant roles in sister chromatid cohesion of the DNA helicase DDX11 and the SMC3 acetyl transferases ESCO1 and ESCO2.PLoS One. 2020 Jan 14;15(1):e0220348. doi: 10.1371/journal.pone.0220348. eCollection 2020. PLoS One. 2020. PMID: 31935221 Free PMC article.
-
Ribosomal biogenesis as an emerging target of neurodevelopmental pathologies.J Neurochem. 2019 Feb;148(3):325-347. doi: 10.1111/jnc.14576. Epub 2018 Nov 12. J Neurochem. 2019. PMID: 30144322 Free PMC article. Review.
-
A novel STAG1 variant associated with congenital clubfoot and microphthalmia: A case report.SAGE Open Med Case Rep. 2024 Aug 31;12:2050313X241277123. doi: 10.1177/2050313X241277123. eCollection 2024. SAGE Open Med Case Rep. 2024. PMID: 39224759 Free PMC article.
References
-
- Michaelis C, Ciosk R, Nasmyth K. Cohesins: chromosomal proteins that prevent premature separation of sister chromatids. Cell 1997, 91:35–45. - PubMed
-
- Onn I, Heidinger‐Pauli JM, Guacci V, Unal E, Koshland DE. Sister chromatid cohesion: a simple concept with a complex reality. Annu Rev Cell Dev Biol 2008, 24:105–129. - PubMed
-
- Gruber S, Haering CH, Nasmyth K. Chromosomal cohesin forms a ring. Cell 2003, 112:765–777. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
