

What is the function of mitochondrial networks? A theoretical assessment of hypotheses and proposal for future research
- PMID: 25847815
- PMCID: PMC4672710
- DOI: 10.1002/bies.201400188
What is the function of mitochondrial networks? A theoretical assessment of hypotheses and proposal for future research
Abstract
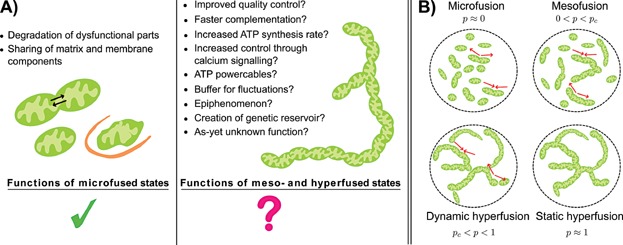
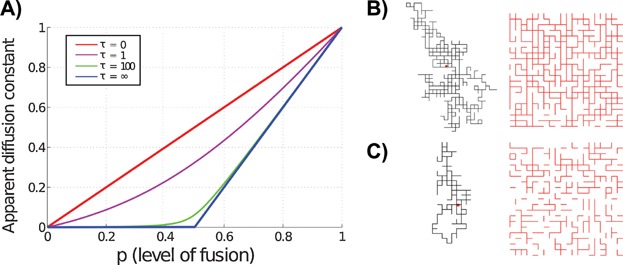
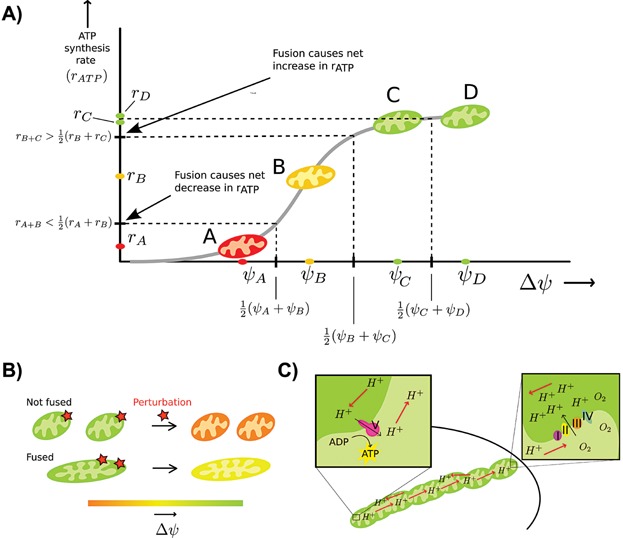
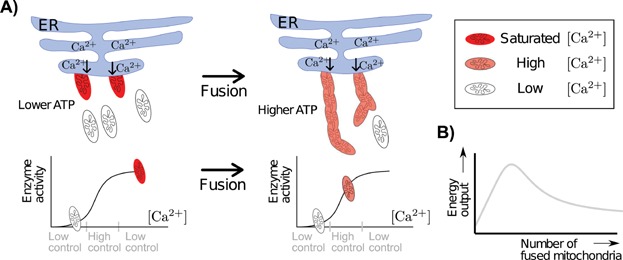
Mitochondria can change their shape from discrete isolated organelles to a large continuous reticulum. The cellular advantages underlying these fused networks are still incompletely understood. In this paper, we describe and compare hypotheses regarding the function of mitochondrial networks. We use mathematical and physical tools both to investigate existing hypotheses and to generate new ones, and we suggest experimental and modelling strategies. Among the novel insights we underline from this work are the possibilities that (i) selective mitophagy is not required for quality control because selective fusion is sufficient; (ii) increased connectivity may have non-linear effects on the diffusion rate of proteins; and (iii) fused networks can act to dampen biochemical fluctuations. We hope to convey to the reader that quantitative approaches can drive advances in the understanding of the physiological advantage of these morphological changes.
Keywords: hypotheses; mathematical biology; mitochondrial dynamics; mitochondrial networks; non-linearities; ultrastructure.
© 2015 The Authors. Bioessays published by WILEY Periodicals, Inc.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
