

Control of bacterial exoelectrogenesis by c-AMP-GMP
- PMID: 25848023
- PMCID: PMC4418907
- DOI: 10.1073/pnas.1419264112
Control of bacterial exoelectrogenesis by c-AMP-GMP
Abstract
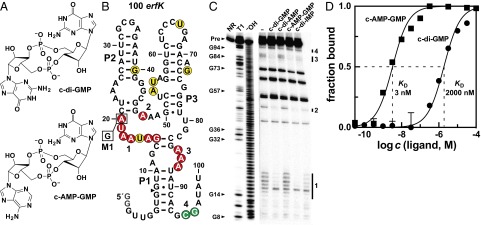
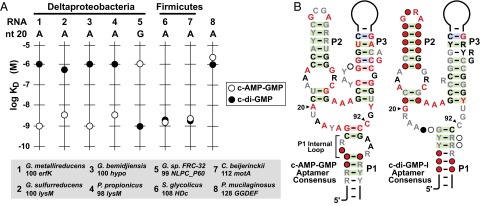
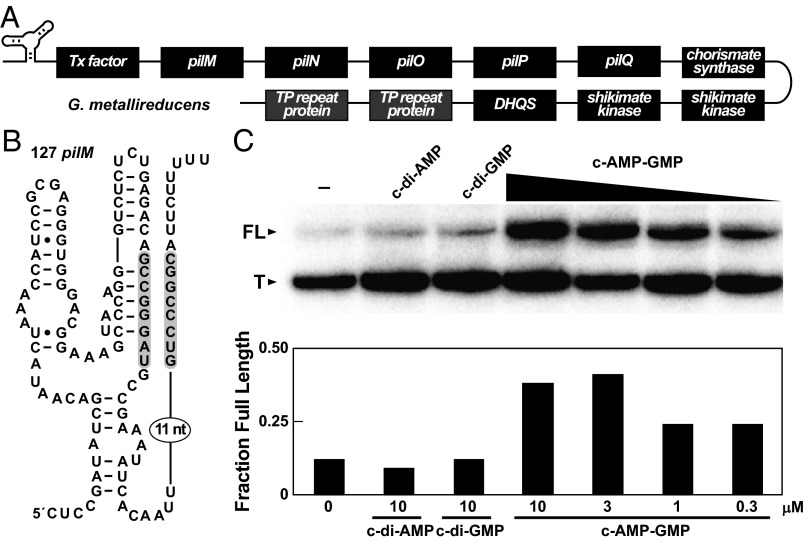
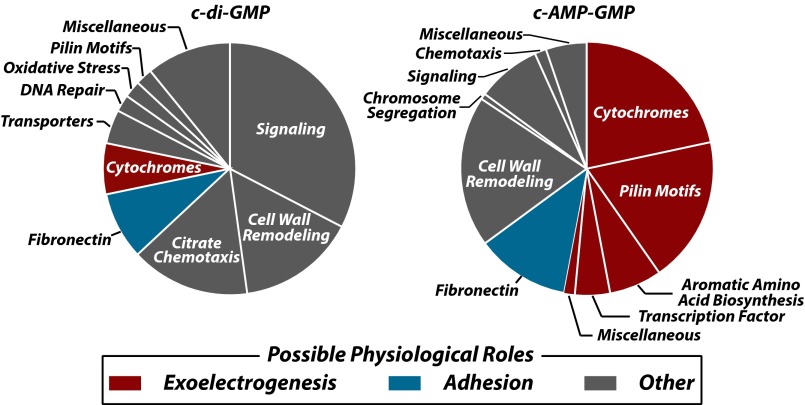
Major changes in bacterial physiology including biofilm and spore formation involve signaling by the cyclic dinucleotides c-di-GMP and c-di-AMP. Recently, another second messenger dinucleotide, c-AMP-GMP, was found to control chemotaxis and colonization by Vibrio cholerae. We have identified a superregulon of genes controlled by c-AMP-GMP in numerous Deltaproteobacteria, including Geobacter species that use extracellular insoluble metal oxides as terminal electron acceptors. This exoelectrogenic process has been studied for its possible utility in energy production and bioremediation. Many genes involved in adhesion, pilin formation, and others that are important for exoelectrogenesis are controlled by members of a variant riboswitch class that selectively bind c-AMP-GMP. These RNAs constitute, to our knowledge, the first known specific receptors for c-AMP-GMP and reveal that this molecule is used by many bacteria to control specialized physiological processes.
Keywords: GEMM; Geobacter; c-di-GMP; riboswitch; second messenger.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Witte G, Hartung S, Büttner K, Hopfner KP. Structural biochemistry of a bacterial checkpoint protein reveals diadenylate cyclase activity regulated by DNA recombination intermediates. Mol Cell. 2008;30(2):167–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
