

Antibody-based screening of cell wall matrix glycans in ferns reveals taxon, tissue and cell-type specific distribution patterns
- PMID: 25848828
- PMCID: PMC4351822
- DOI: 10.1186/s12870-014-0362-8
Antibody-based screening of cell wall matrix glycans in ferns reveals taxon, tissue and cell-type specific distribution patterns
Abstract
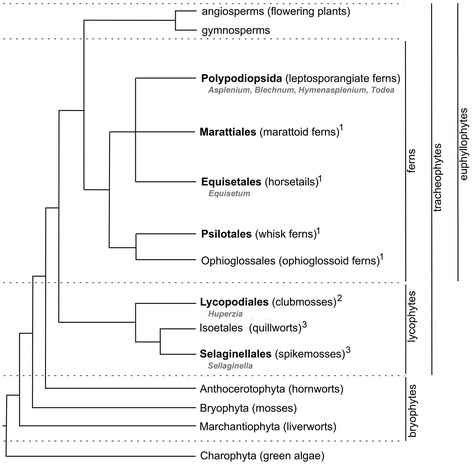
Background: While it is kno3wn that complex tissues with specialized functions emerged during land plant evolution, it is not clear how cell wall polymers and their structural variants are associated with specific tissues or cell types. Moreover, due to the economic importance of many flowering plants, ferns have been largely neglected in cell wall comparative studies.
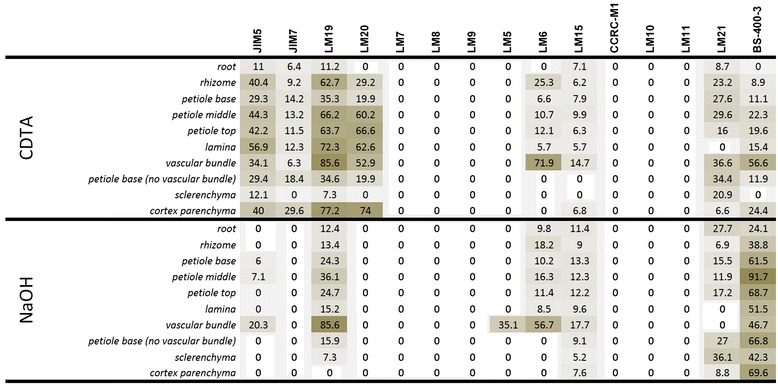
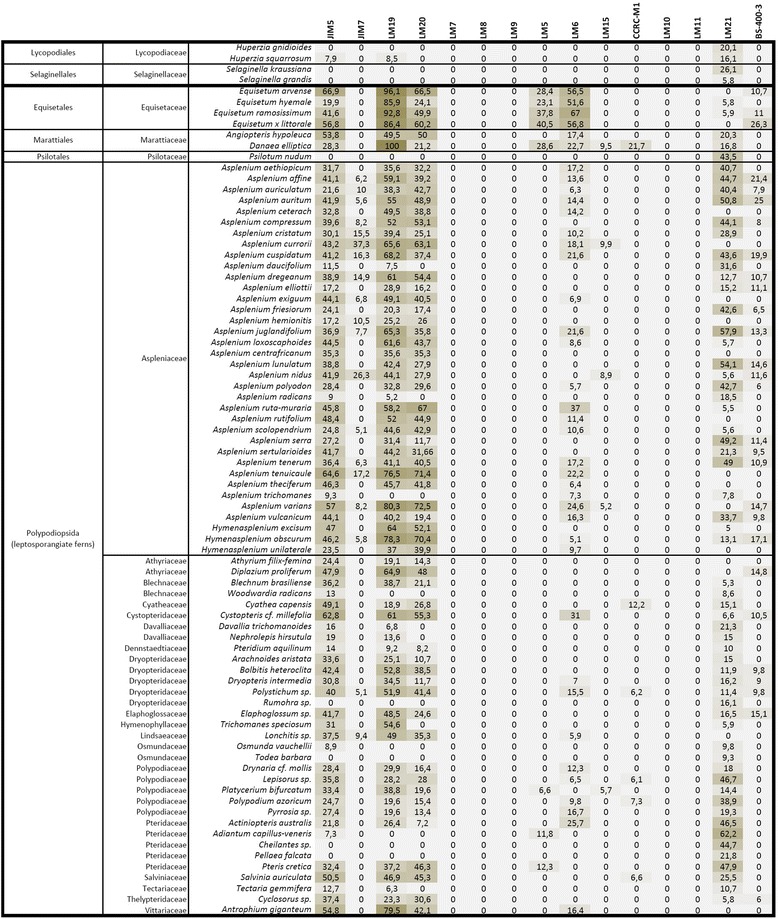
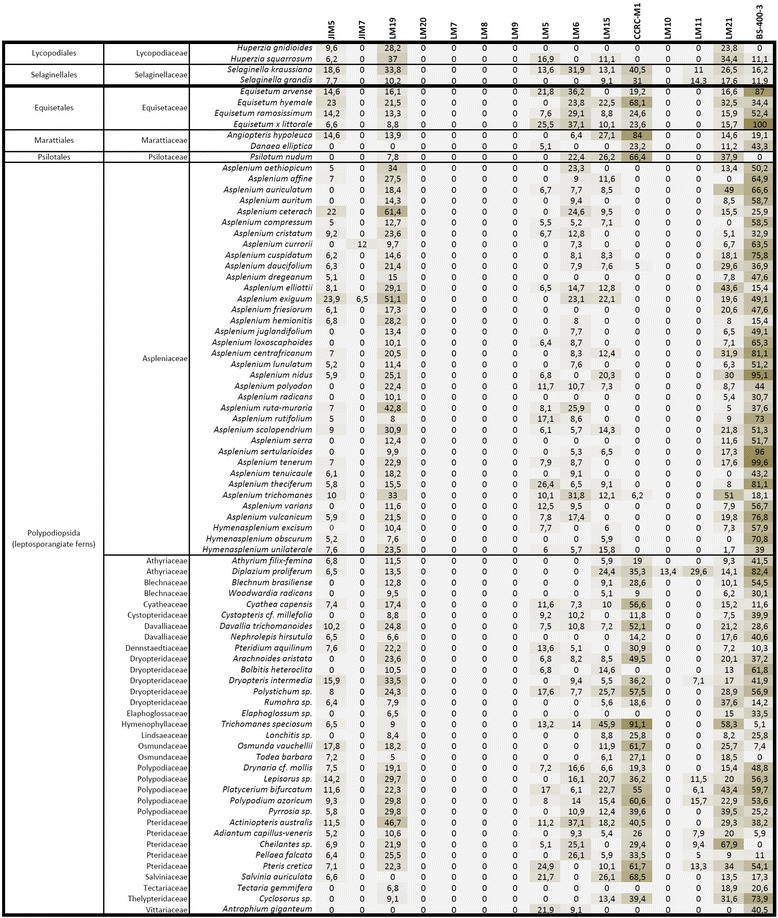
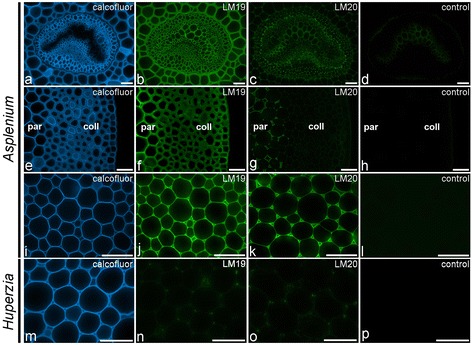
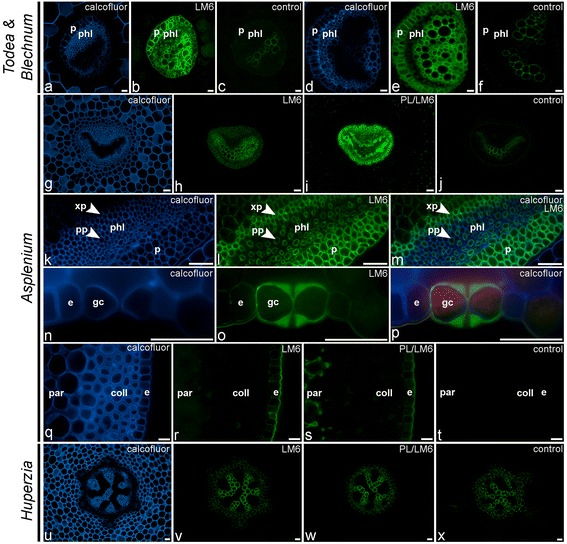
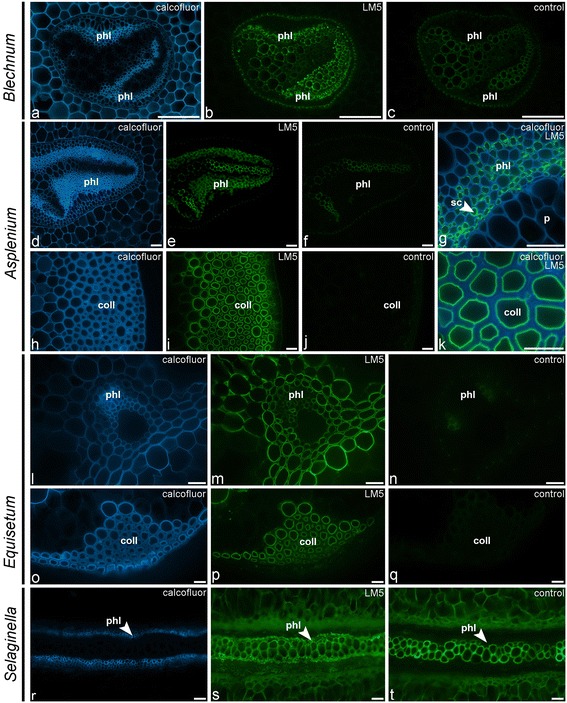
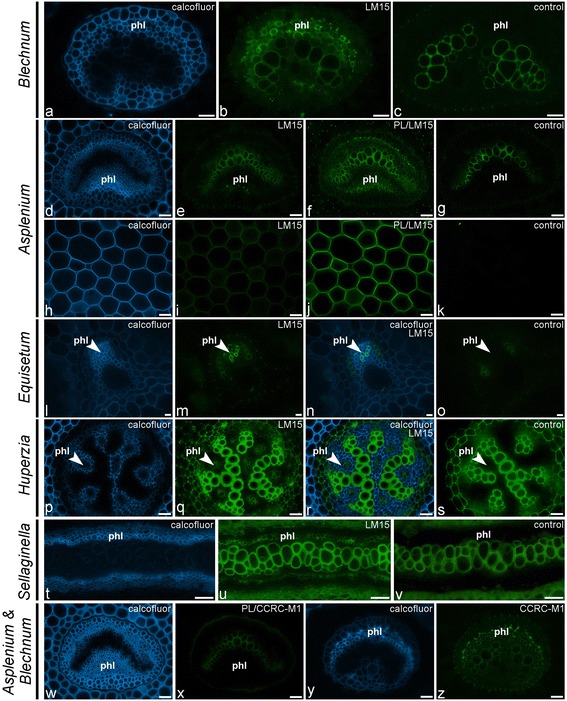
Results: To explore fern cell wall diversity sets of monoclonal antibodies directed to matrix glycans of angiosperm cell walls have been used in glycan microarray and in situ analyses with 76 fern species and four species of lycophytes. All major matrix glycans were present as indicated by epitope detection with some variations in abundance. Pectic HG epitopes were of low abundance in lycophytes and the CCRC-M1 fucosylated xyloglucan epitope was largely absent from the Aspleniaceae. The LM15 XXXG epitope was detected widely across the ferns and specifically associated with phloem cell walls and similarly the LM11 xylan epitope was associated with xylem cell walls. The LM5 galactan and LM6 arabinan epitopes, linked to pectic supramolecules in angiosperms, were associated with vascular structures with only limited detection in ground tissues. Mannan epitopes were found to be associated with the development of mechanical tissues. We provided the first evidence for the presence of MLG in leptosporangiate ferns.
Conclusions: The data sets indicate that cell wall diversity in land plants is multifaceted and that matrix glycan epitopes display complex spatio-temporal and phylogenetic distribution patterns that are likely to relate to the evolution of land plant body plans.
Figures
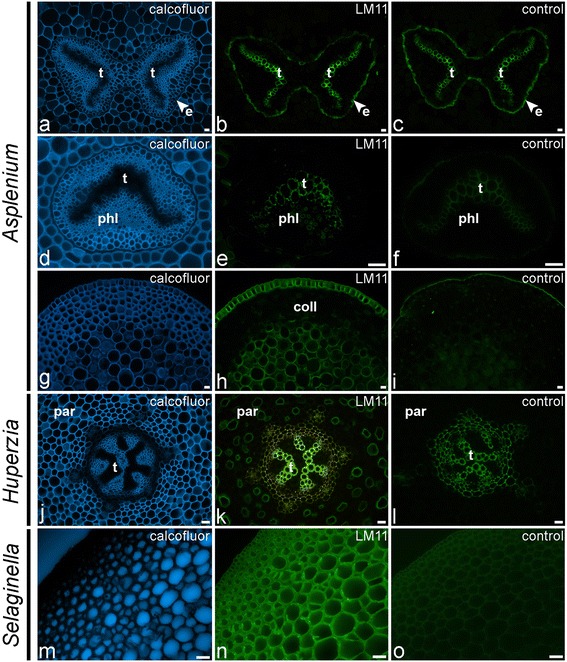
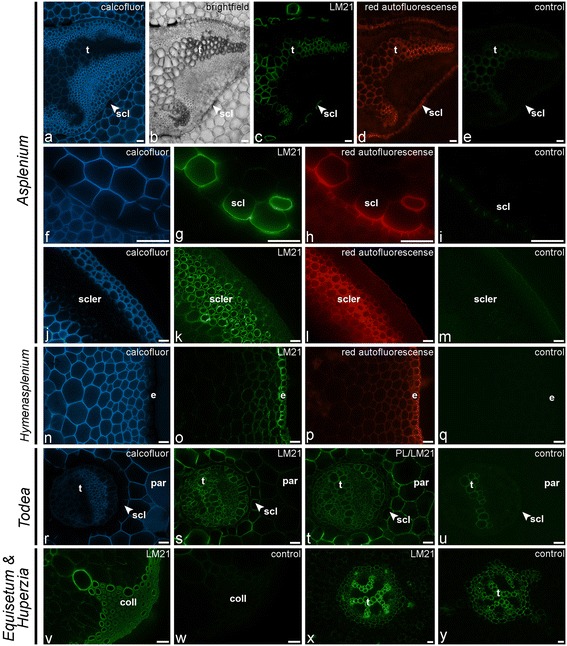

References
-
- Bateman RM, Crane PR, SiMichele WA, Kenrick PR, Rowe NP, Speck T, Stein EW. Early evolution of land plants: phylogeny, physiology, and ecology of the primary terrestrial radiation. Ann Rev Ecol System. 1998;29:263–292. doi: 10.1146/annurev.ecolsys.29.1.263. - DOI
-
- Evert RF. Esau's plant anatomy. Meristems, cells, and tissues of the plant body: their structure, function, and development. New Jersey: John Wiley and Sons, Inc; 2006.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
