

Proteolysis of virulence regulator ToxR is associated with entry of Vibrio cholerae into a dormant state
- PMID: 25849031
- PMCID: PMC4388833
- DOI: 10.1371/journal.pgen.1005145
Proteolysis of virulence regulator ToxR is associated with entry of Vibrio cholerae into a dormant state
Abstract
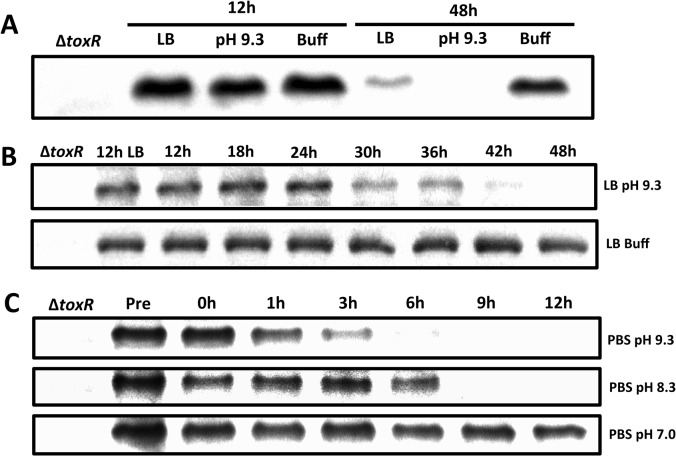
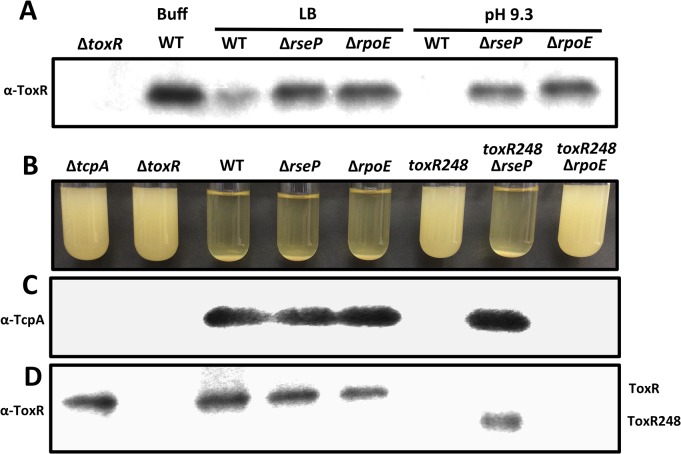
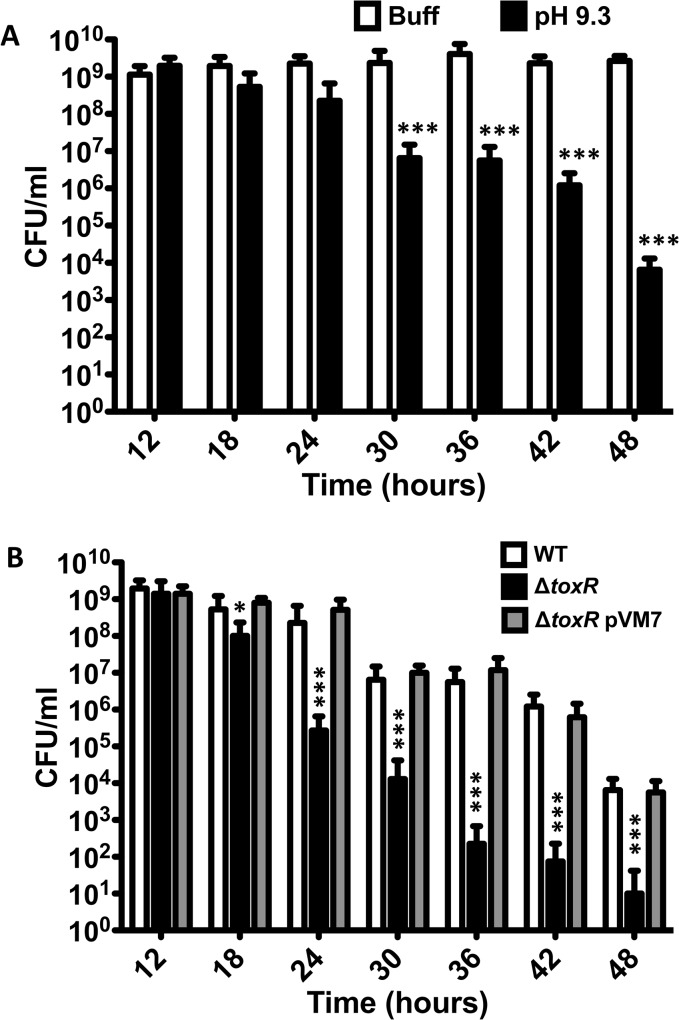
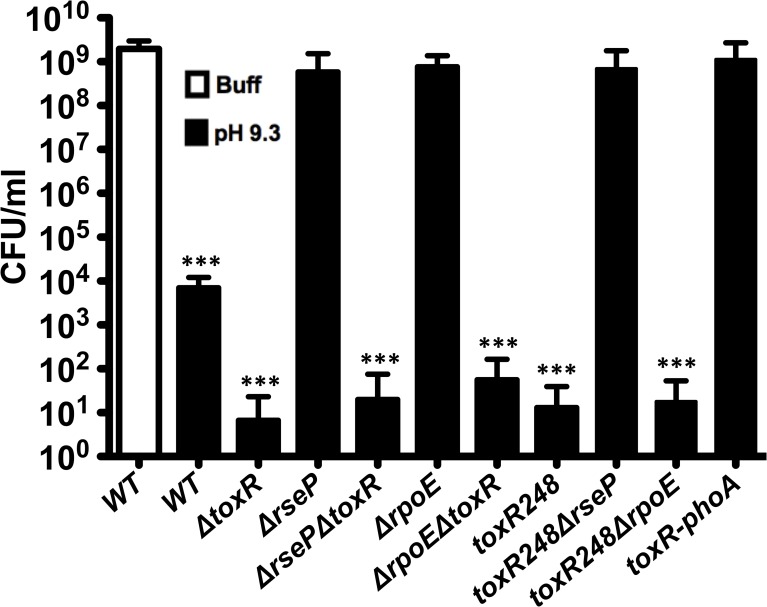
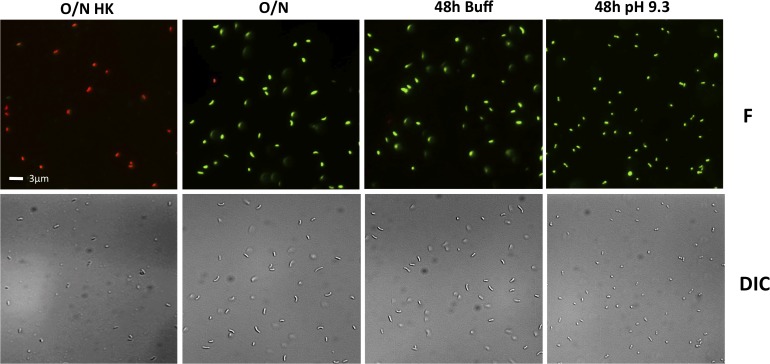
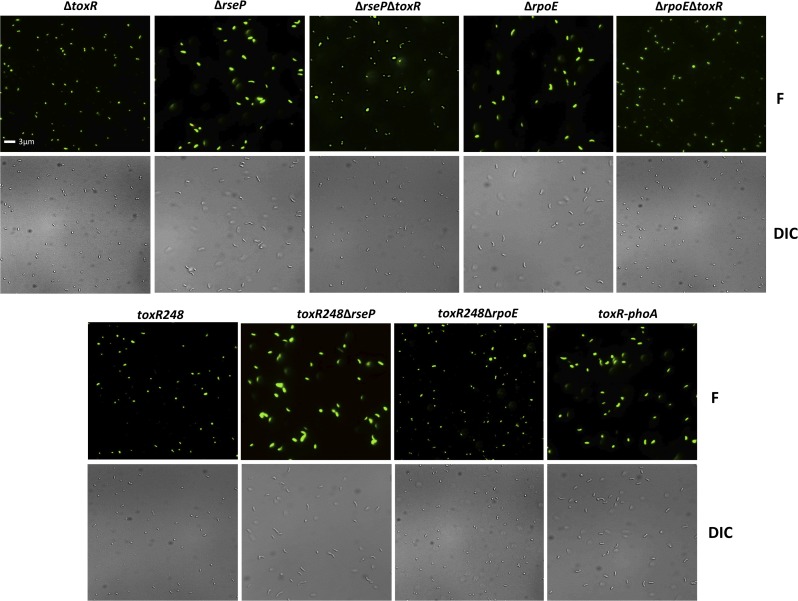
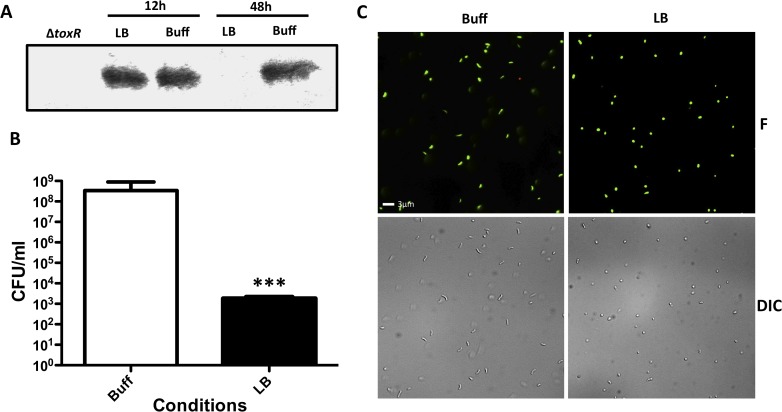
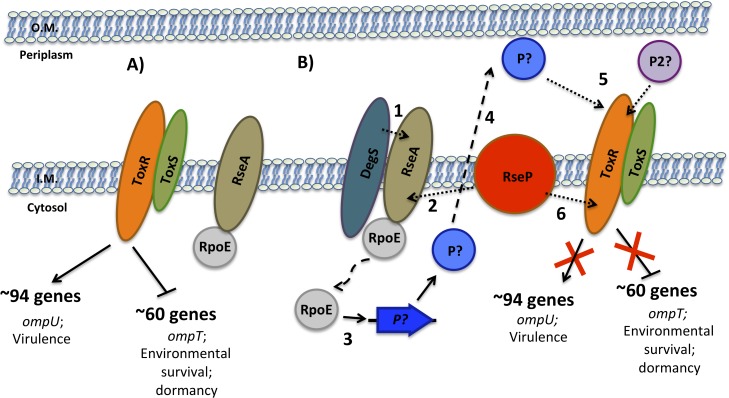
Vibrio cholerae O1 is a natural inhabitant of aquatic environments and causes the diarrheal disease, cholera. Two of its primary virulence regulators, TcpP and ToxR, are localized in the inner membrane. TcpP is encoded on the Vibrio Pathogenicity Island (VPI), a horizontally acquired mobile genetic element, and functions primarily in virulence gene regulation. TcpP has been shown to undergo regulated intramembrane proteolysis (RIP) in response to environmental conditions that are unfavorable for virulence gene expression. ToxR is encoded in the ancestral genome and is present in non-pathogenic strains of V. cholerae, indicating it has roles outside of the human host. In this study, we show that ToxR undergoes RIP in V. cholerae in response to nutrient limitation at alkaline pH, a condition that occurs during the stationary phase of growth. This process involves the site-2 protease RseP (YaeL), and is dependent upon the RpoE-mediated periplasmic stress response, as deletion mutants for the genes encoding these two proteins cannot proteolyze ToxR under nutrient limitation at alkaline pH. We determined that the loss of ToxR, genetically or by proteolysis, is associated with entry of V. cholerae into a dormant state in which the bacterium is normally found in the aquatic environment called viable but nonculturable (VBNC). Strains that can proteolyze ToxR, or do not encode it, lose culturability, experience a change in morphology associated with cells in VBNC, yet remain viable under nutrient limitation at alkaline pH. On the other hand, mutant strains that cannot proteolyze ToxR remain culturable and maintain the morphology of cells in an active state of growth. Overall, our findings provide a link between the proteolysis of a virulence regulator and the entry of a pathogen into an environmentally persistent state.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Almagro-Moreno S, Taylor RK (2013) Cholera: Environmental Reservoirs and Impact on Disease Transmission. Microbiol Spectrum 1(2):OH-0003-2012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
