

Structural and functional characterization of TesB from Yersinia pestis reveals a unique octameric arrangement of hotdog domains
- PMID: 25849407
- PMCID: PMC4388271
- DOI: 10.1107/S1399004715002527
Structural and functional characterization of TesB from Yersinia pestis reveals a unique octameric arrangement of hotdog domains
Abstract
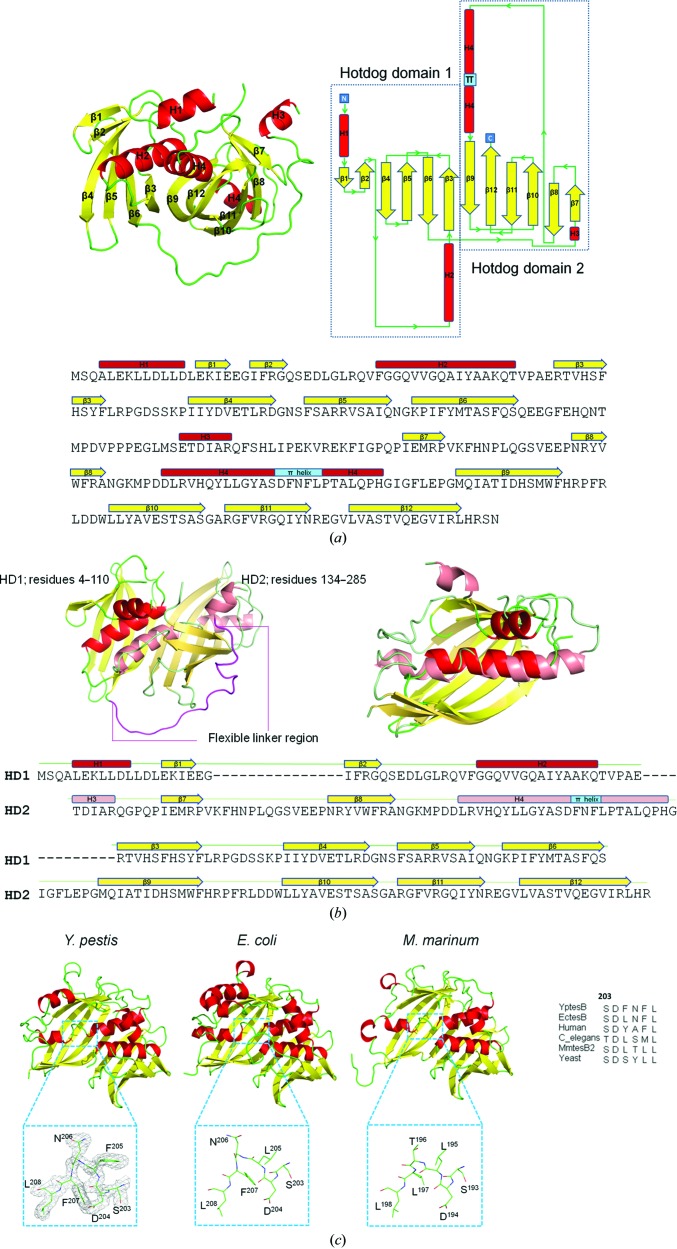
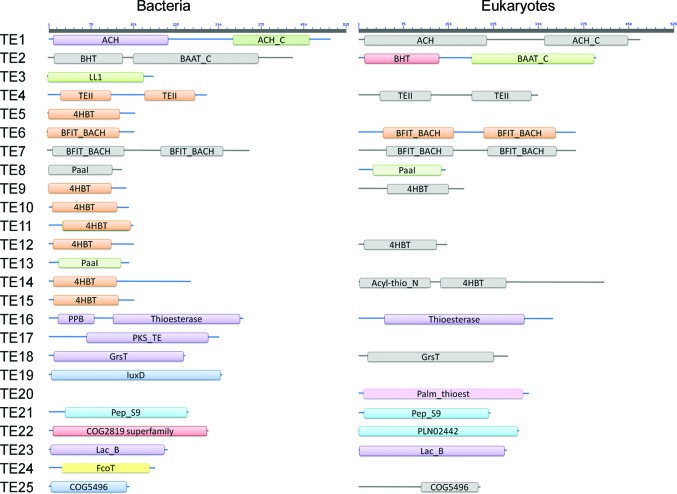
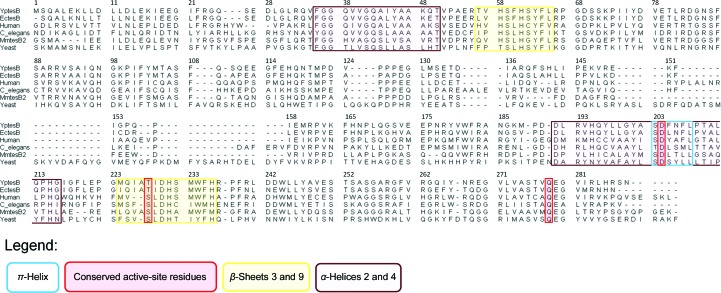
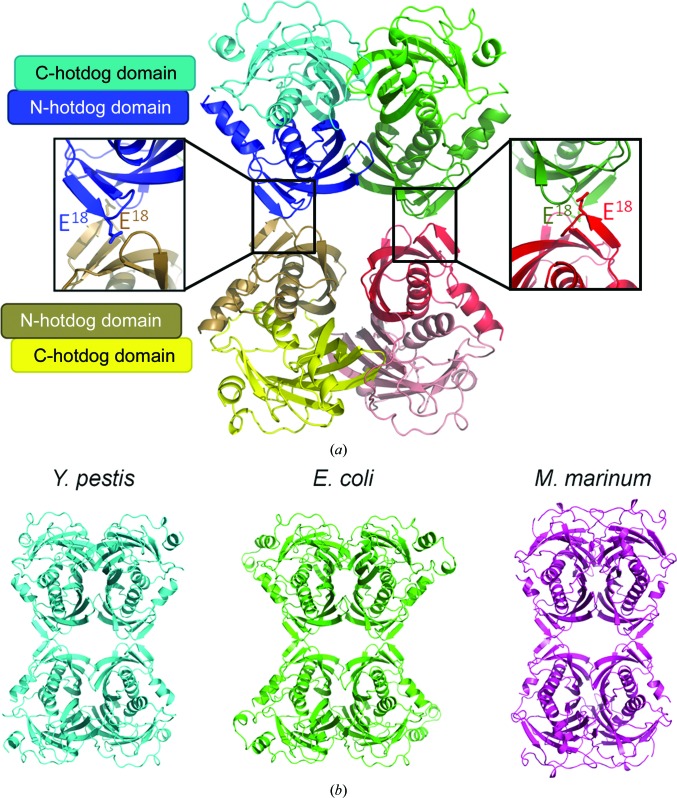
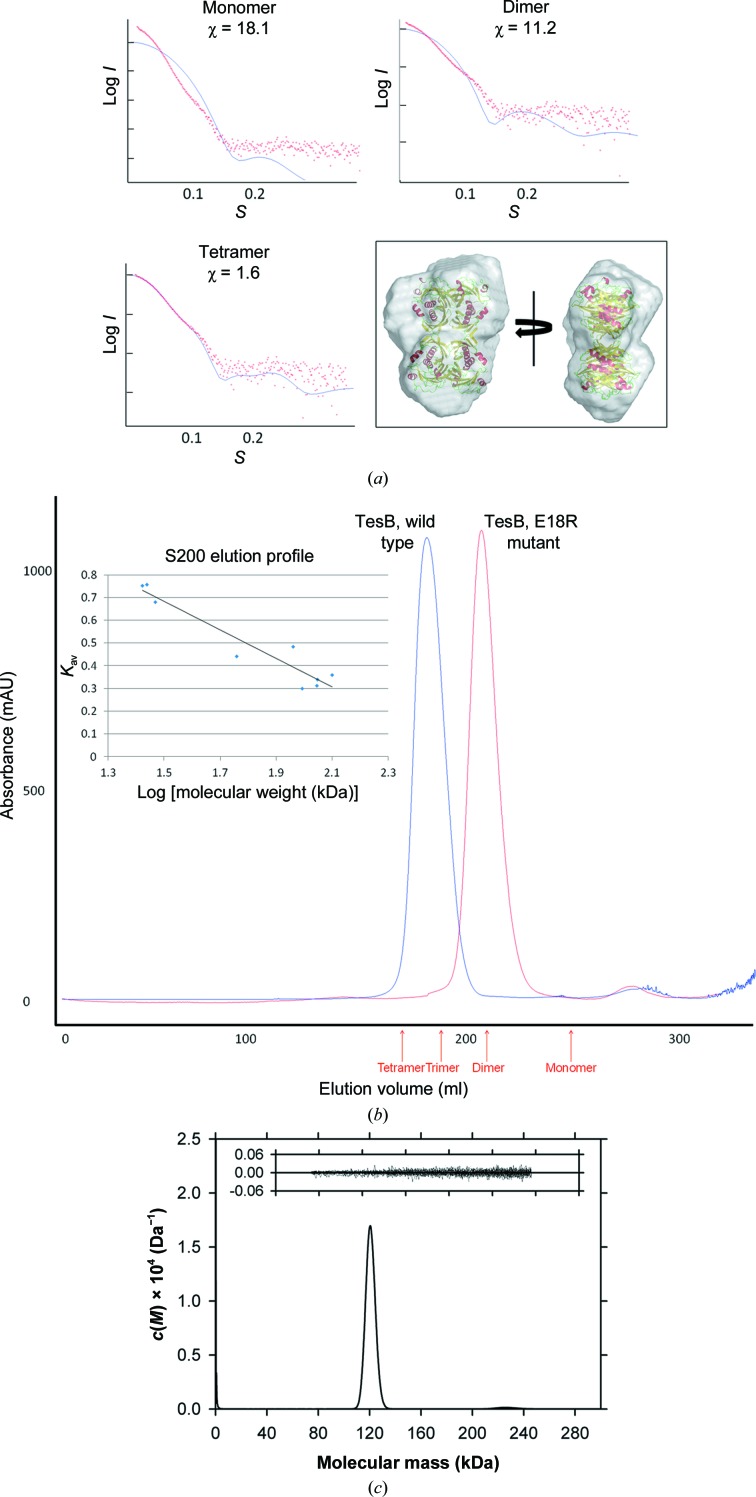
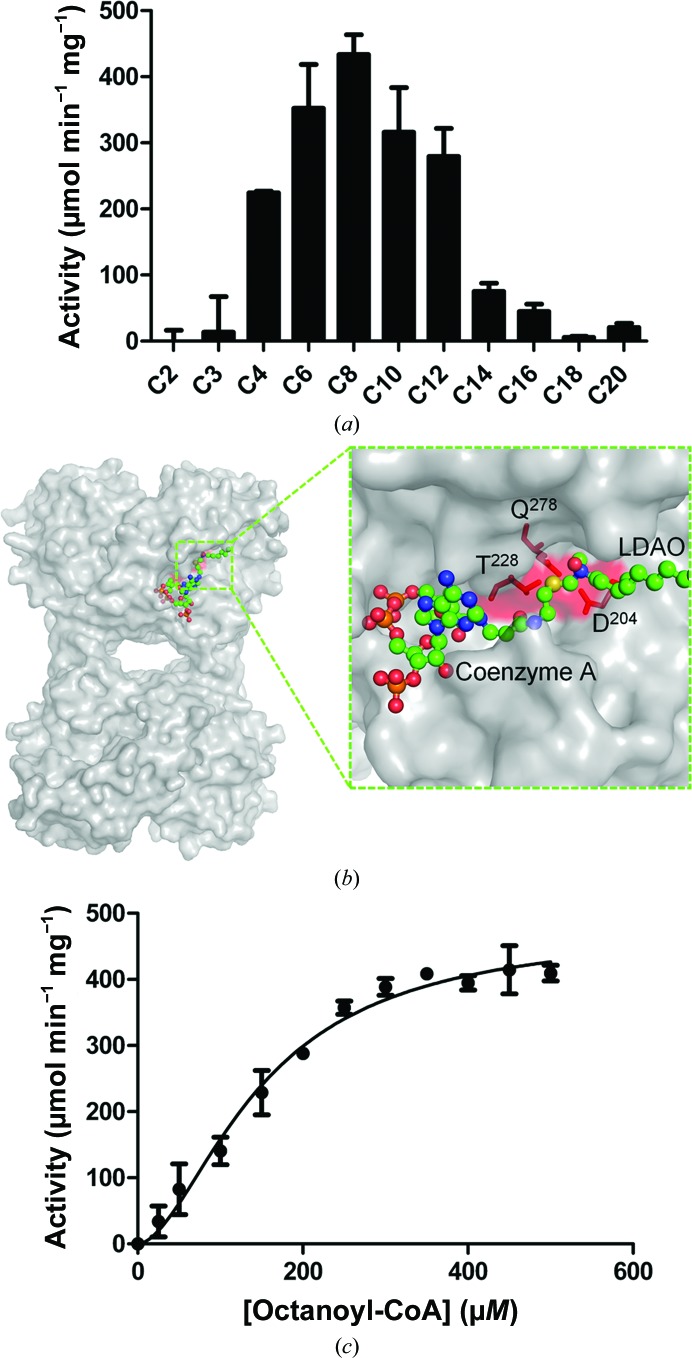
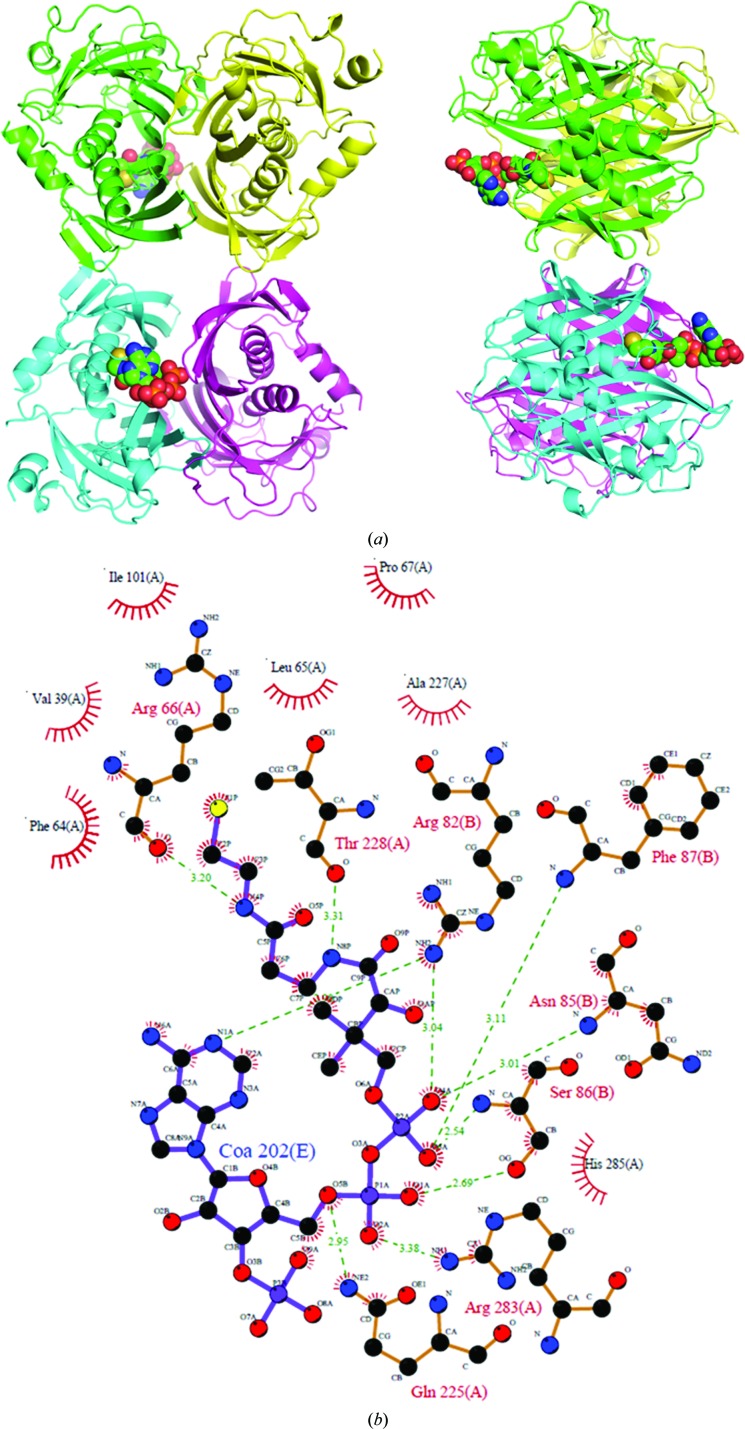
Acyl-CoA thioesterases catalyse the hydrolysis of the thioester bonds present within a wide range of acyl-CoA substrates, releasing free CoASH and the corresponding fatty-acyl conjugate. The TesB-type thioesterases are members of the TE4 thioesterase family, one of 25 thioesterase enzyme families characterized to date, and contain two fused hotdog domains in both prokaryote and eukaryote homologues. Only two structures have been elucidated within this enzyme family, and much of the current understanding of the TesB thioesterases has been based on the Escherichia coli structure. Yersinia pestis, a highly virulent bacterium, encodes only one TesB-type thioesterase in its genome; here, the structural and functional characterization of this enzyme are reported, revealing unique elements both within the protomer and quaternary arrangements of the hotdog domains which have not been reported previously in any thioesterase family. The quaternary structure, confirmed using a range of structural and biophysical techniques including crystallography, small-angle X-ray scattering, analytical ultracentrifugation and size-exclusion chromatography, exhibits a unique octameric arrangement of hotdog domains. Interestingly, the same biological unit appears to be present in both TesB structures solved to date, and is likely to be a conserved and distinguishing feature of TesB-type thioesterases. Analysis of the Y. pestis TesB thioesterase activity revealed a strong preference for octanoyl-CoA and this is supported by structural analysis of the active site. Overall, the results provide novel insights into the structure of TesB thioesterases which are likely to be conserved and distinguishing features of the TE4 thioesterase family.
Keywords: TesB; Yersinia pestis; acyl-CoA thioesterases.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
