

An analysis of critical factors for quantitative immunoblotting
- PMID: 25852189
- PMCID: PMC4401487
- DOI: 10.1126/scisignal.2005966
An analysis of critical factors for quantitative immunoblotting
Abstract
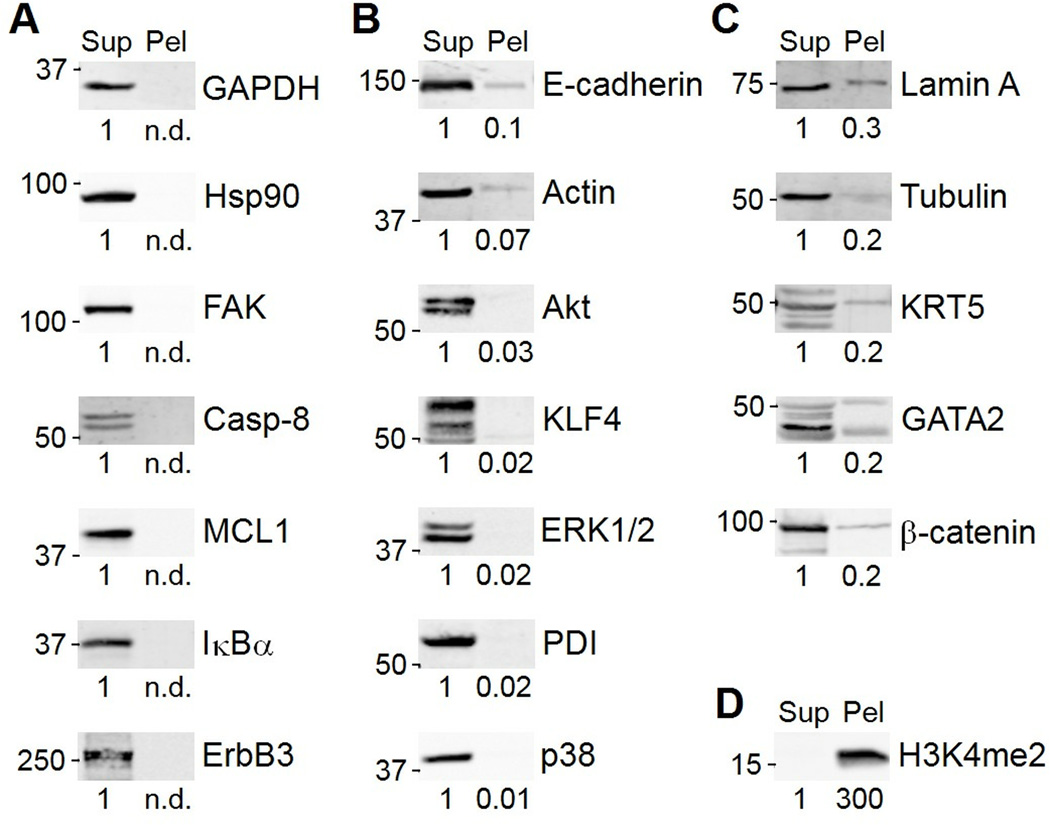
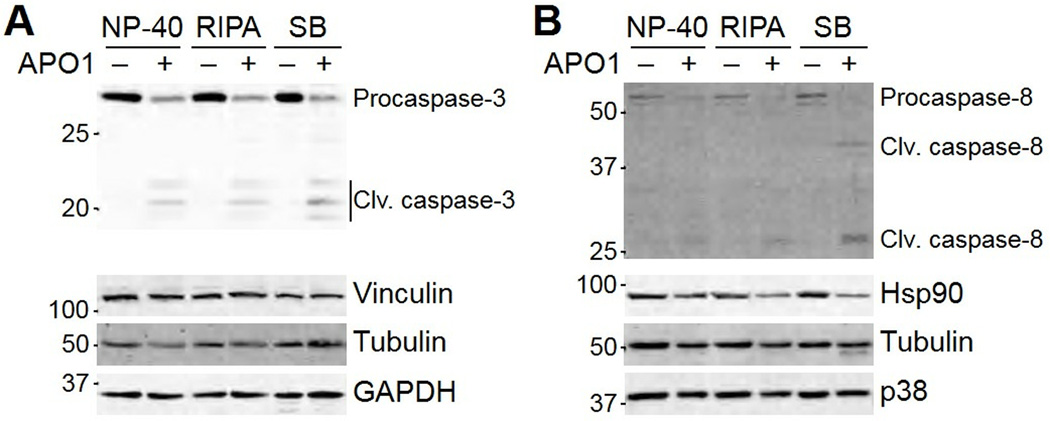
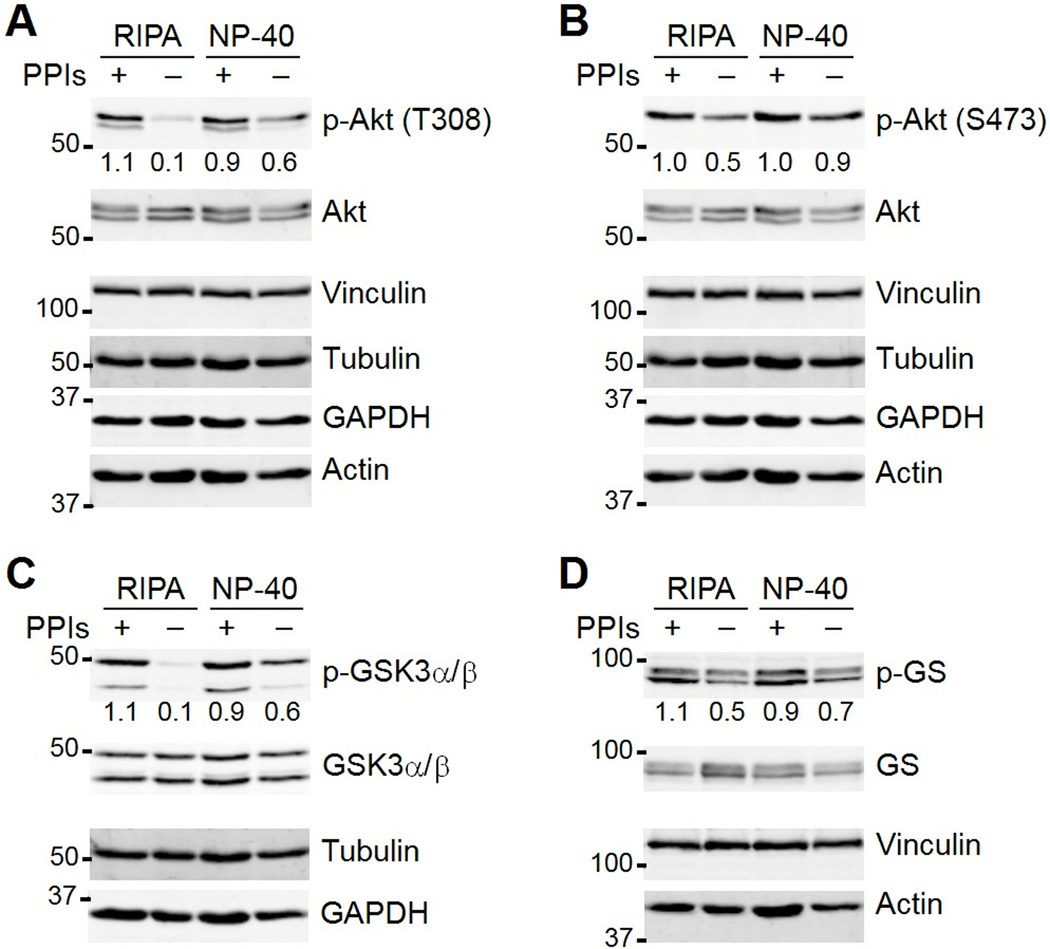
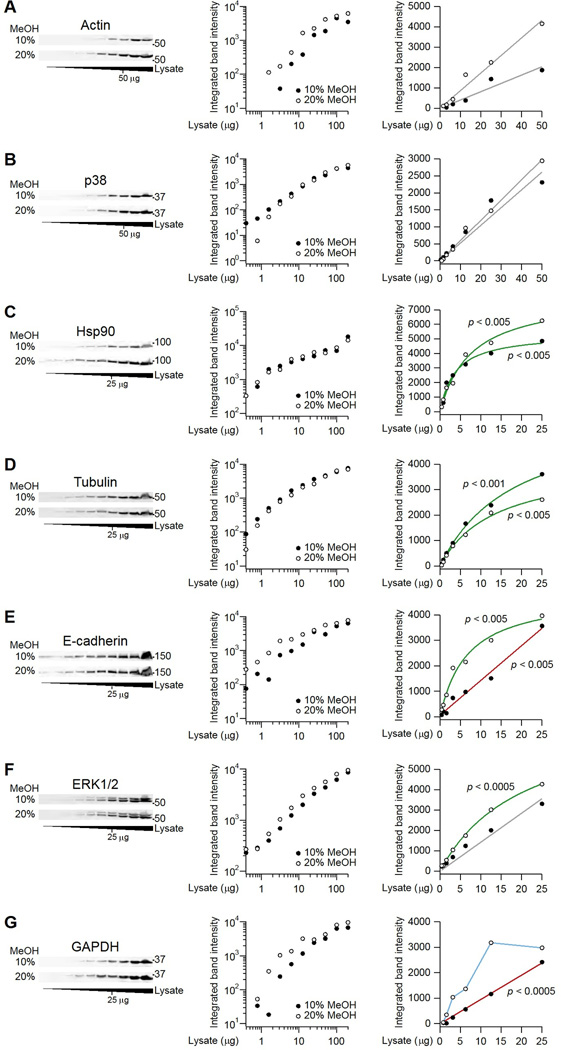
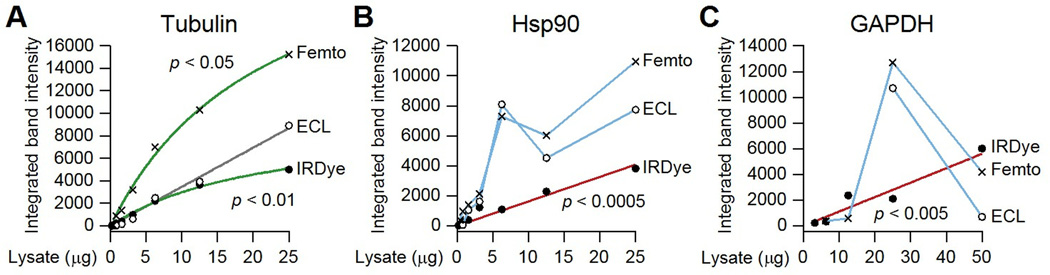
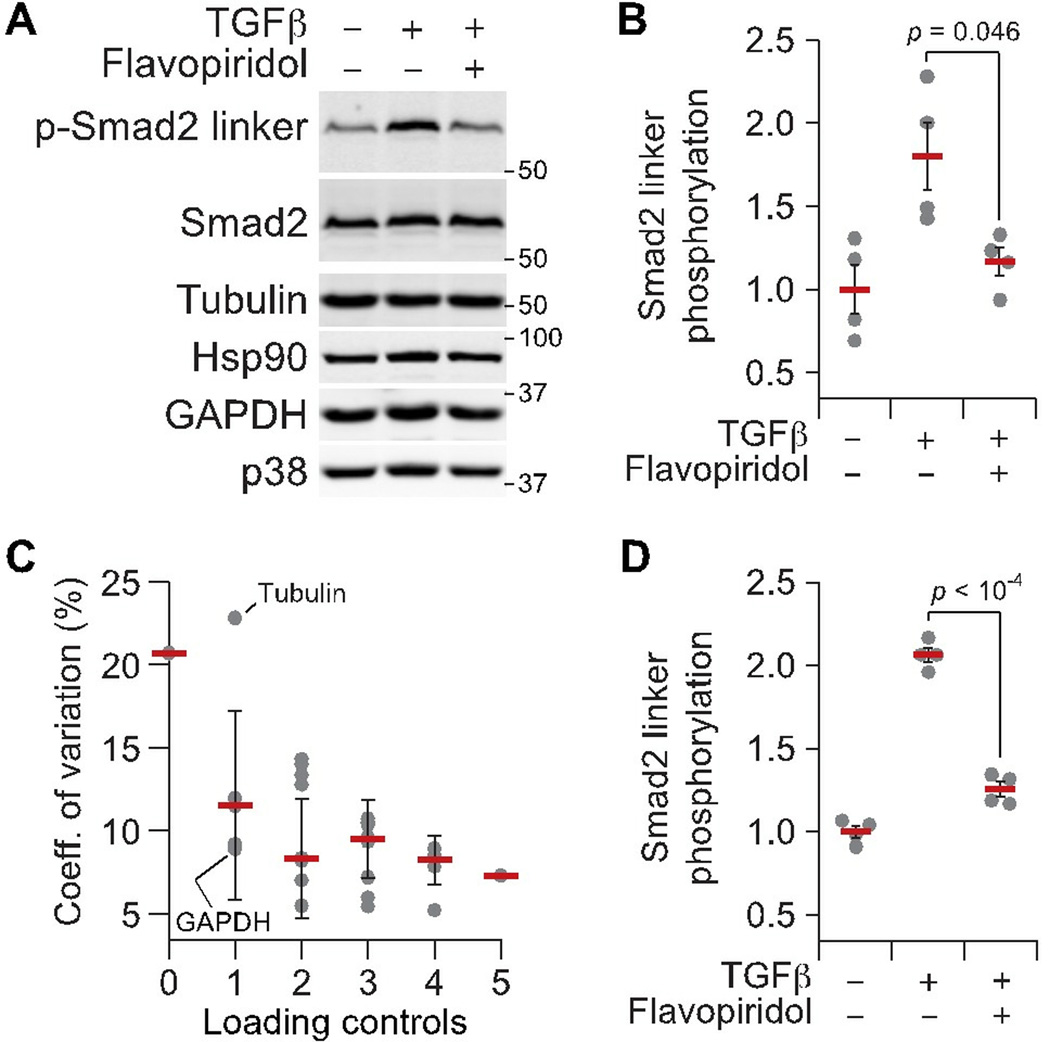
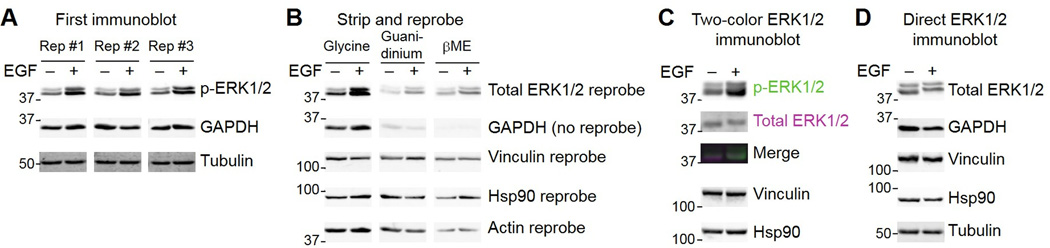
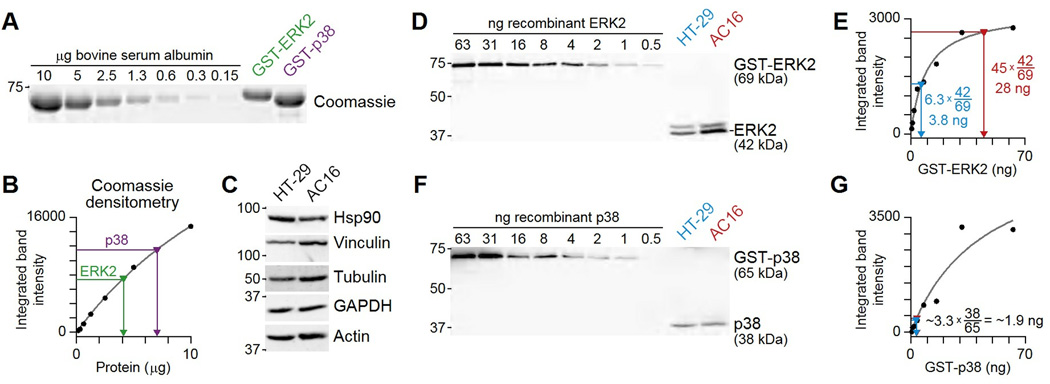
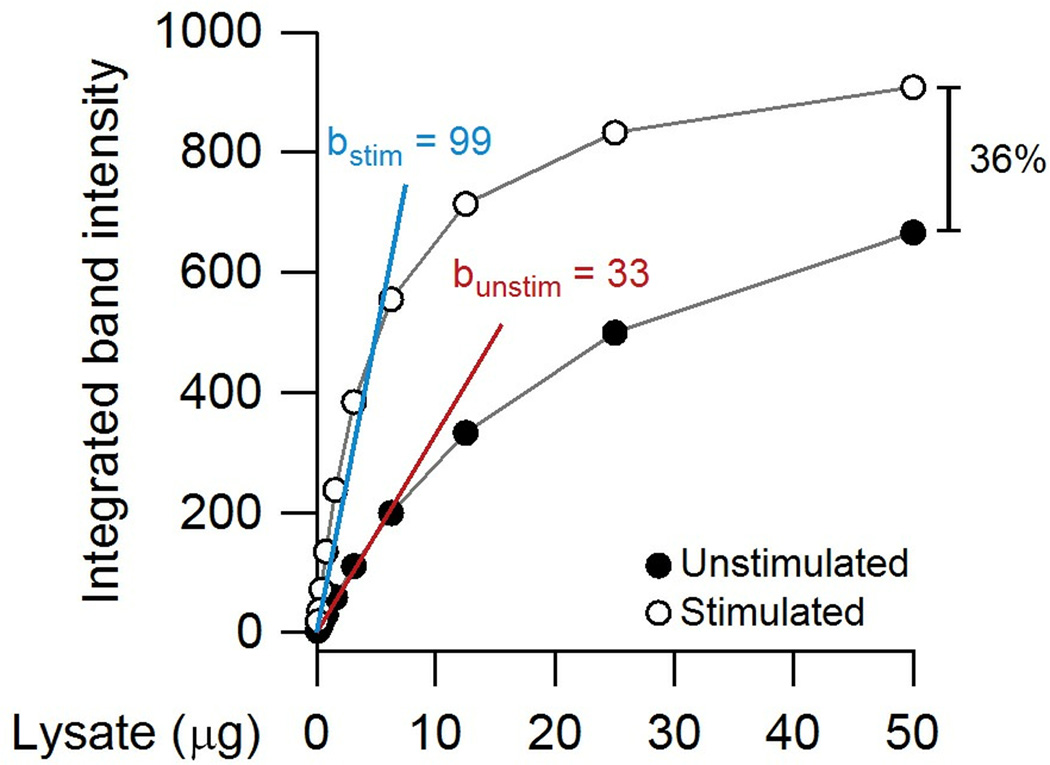
Immunoblotting (also known as Western blotting) combined with digital image analysis can be a reliable method for analyzing the abundance of proteins and protein modifications, but not every immunoblot-analysis combination produces an accurate result. I illustrate how sample preparation, protocol implementation, detection scheme, and normalization approach profoundly affect the quantitative performance of immunoblotting. This study implemented diagnostic experiments that assess an immunoblot-analysis workflow for accuracy and precision. The results showed that ignoring such diagnostics can lead to pseudoquantitative immunoblot data that markedly overestimate or underestimate true differences in protein abundance.
Copyright © 2015, American Association for the Advancement of Science.
Figures
References
-
- Green MR, Sambrook J. Molecular cloning: a laboratory manual. ed. 4th. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2012. p. 1890.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
