

Differential modulation of repetitive firing and synchronous network activity in neocortical interneurons by inhibition of A-type K(+) channels and Ih
- PMID: 25852481
- PMCID: PMC4364302
- DOI: 10.3389/fncel.2015.00089
Differential modulation of repetitive firing and synchronous network activity in neocortical interneurons by inhibition of A-type K(+) channels and Ih
Abstract
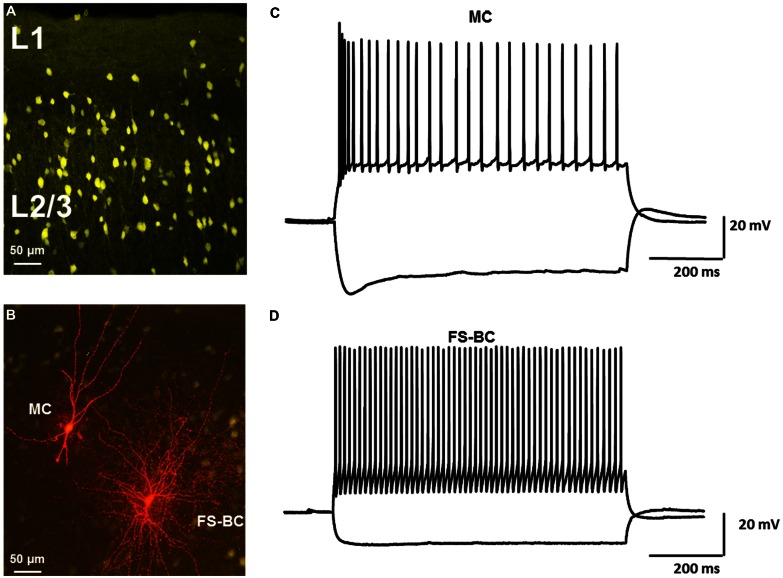
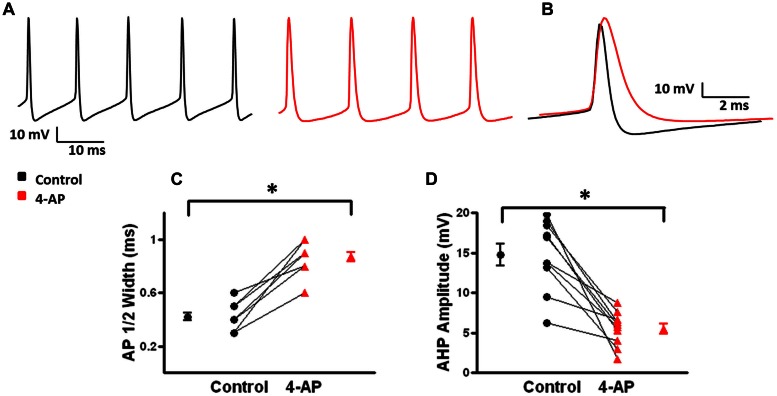
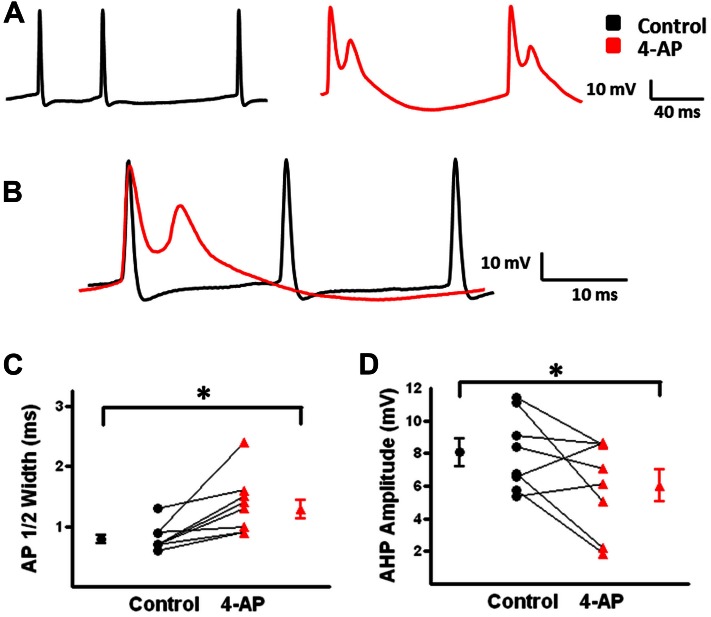
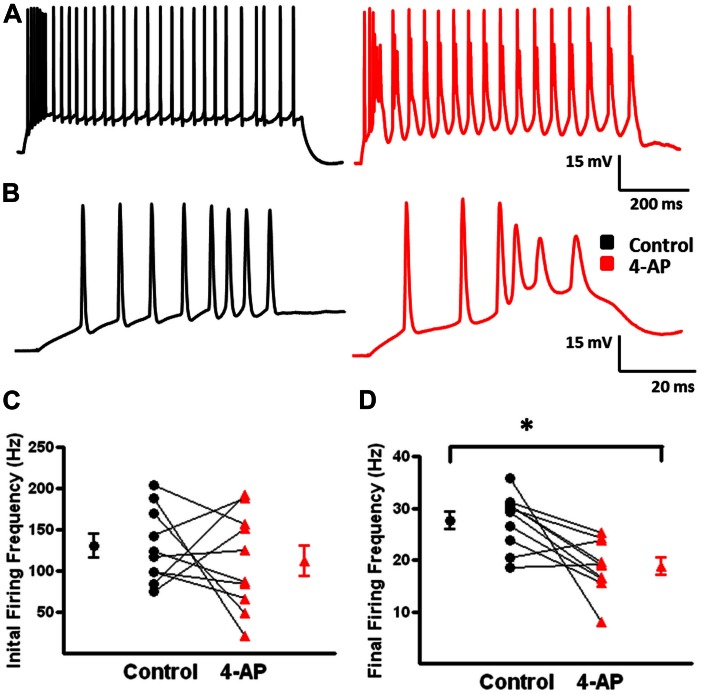
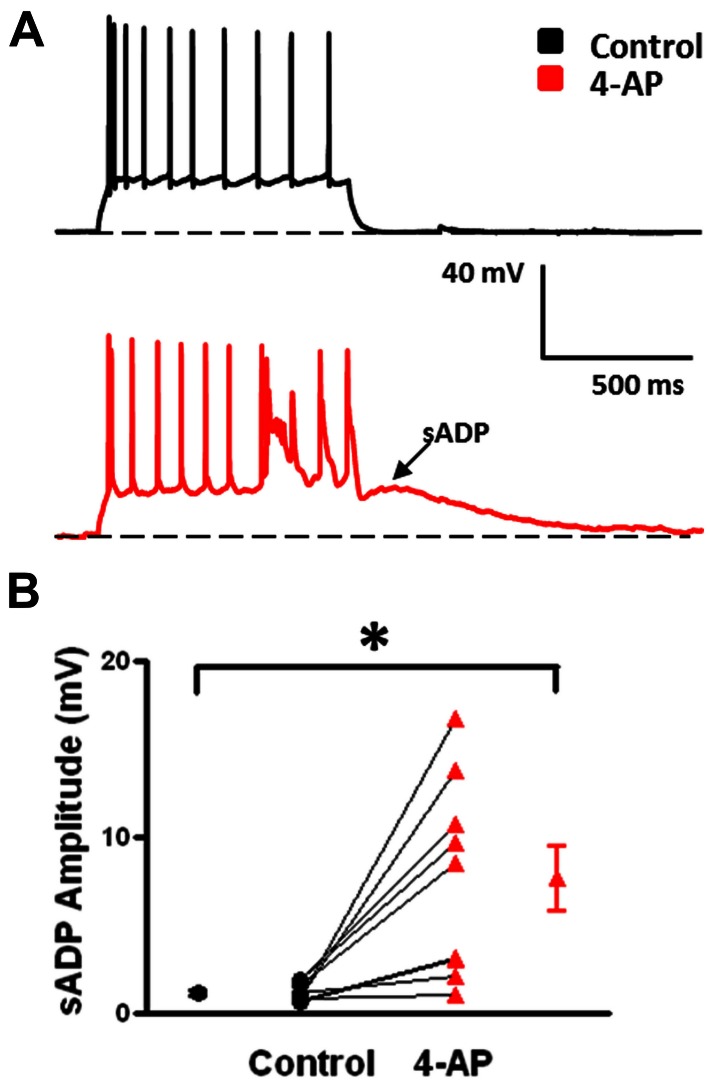
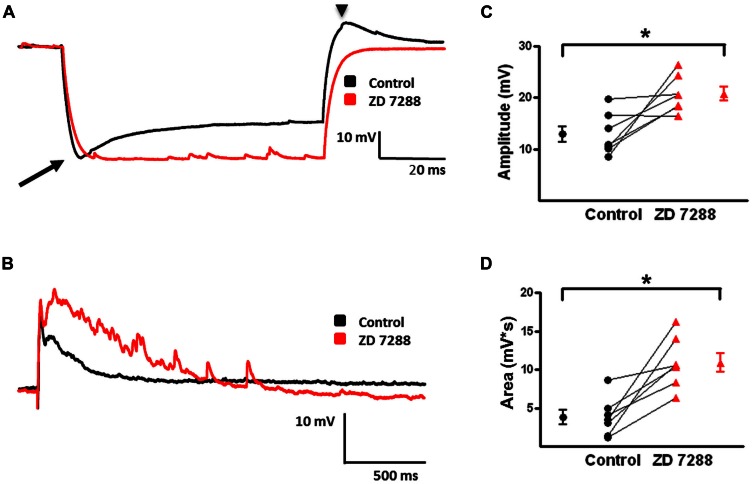
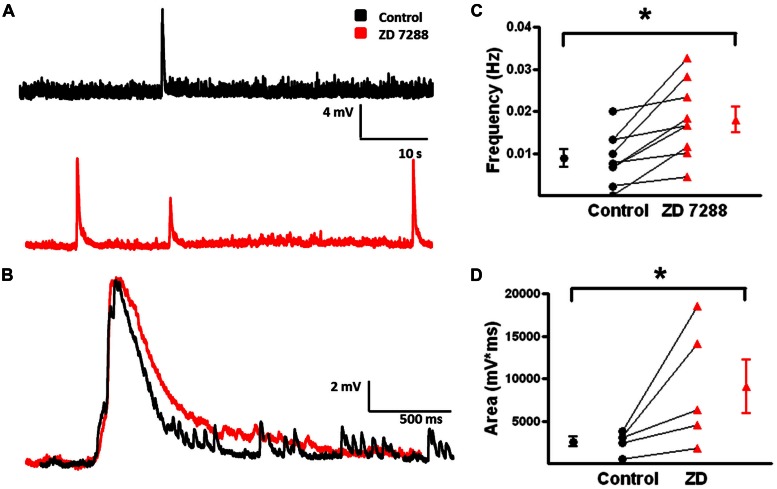
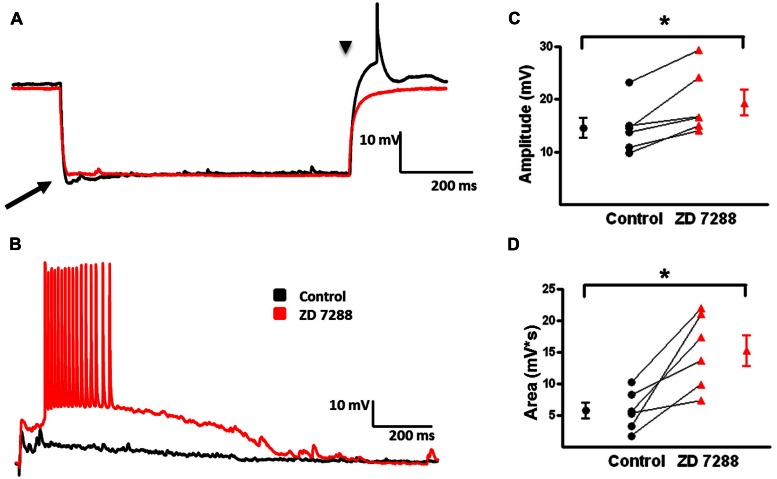
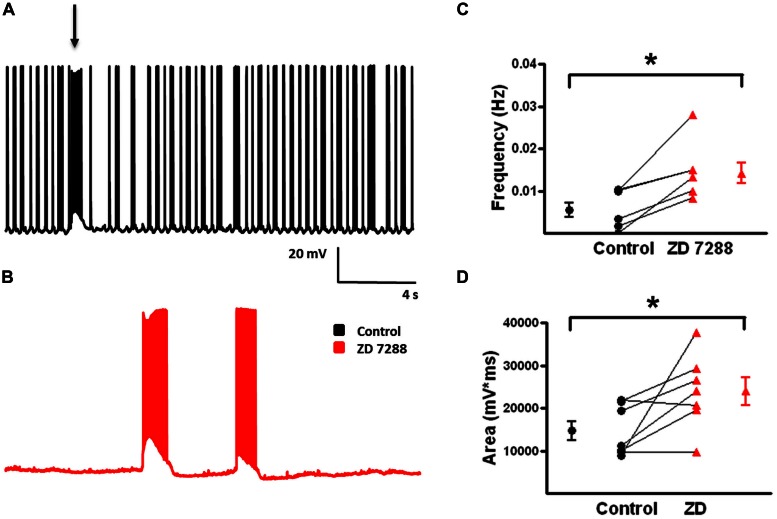
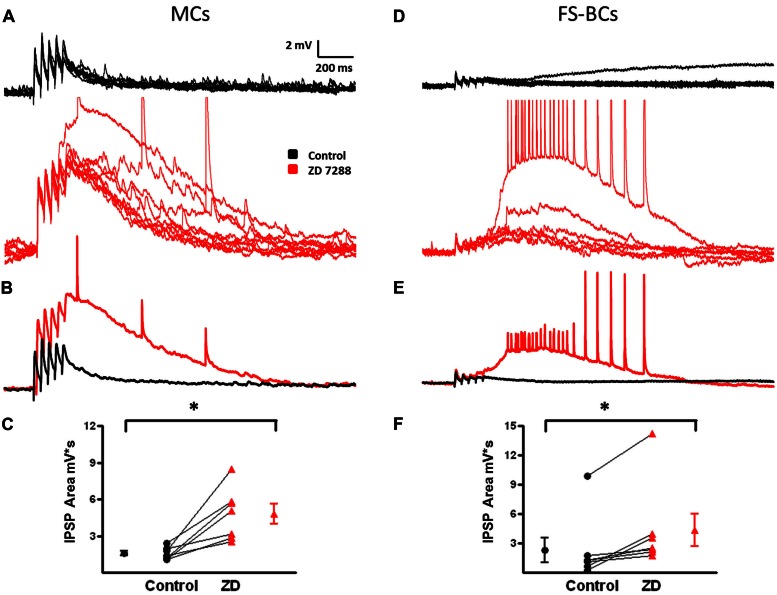
GABAergic interneurons provide the main source of inhibition in the neocortex and are important in regulating neocortical network activity. In the presence 4-aminopyridine (4-AP), CNQX, and D-APV, large amplitude GABAA-receptor mediated depolarizing responses were observed in the neocortex. GABAergic networks are comprised of several types of interneurons, each with its own protein expression pattern, firing properties, and inhibitory role in network activity. Voltage-gated ion channels, especially A-type K(+) channels, differentially regulate passive membrane properties, action potential (AP) waveform, and repetitive firing properties in interneurons depending on their composition and localization. HCN channels are known modulators of pyramidal cell intrinsic excitability and excitatory network activity. Little information is available regarding how HCN channels functionally modulate excitability of individual interneurons and inhibitory networks. In this study, we examined the effect of 4-AP on intrinsic excitability of fast-spiking basket cells (FS-BCs) and Martinotti cells (MCs). 4-AP increased the duration of APs in both FS-BCs and MCs. The repetitive firing properties of MCs were differentially affected compared to FS-BCs. We also examined the effect of Ih inhibition on synchronous GABAergic depolarizations and synaptic integration of depolarizing IPSPs. ZD 7288 enhanced the amplitude and area of evoked GABAergic responses in both cell types. Similarly, the frequency and area of spontaneous GABAergic depolarizations in both FS-BCs and MCs were increased in presence of ZD 7288. Synaptic integration of IPSPs in MCs was significantly enhanced, but remained unaltered in FS-BCs. These results indicate that 4-AP differentially alters the firing properties of interneurons, suggesting MCs and FS-BCs may have unique roles in GABAergic network synchronization. Enhancement of GABAergic network synchronization by ZD 7288 suggests that HCN channels attenuate inhibitory network activity.
Keywords: 4-AP; A-type K+ channels; GABAergic interneurons; HCN channels; Ih; Martinotti cells; basket cells; neocortex; synchronization.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous