

Researching glutamate - induced cytotoxicity in different cell lines: a comparative/collective analysis/study
- PMID: 25852482
- PMCID: PMC4362409
- DOI: 10.3389/fncel.2015.00091
Researching glutamate - induced cytotoxicity in different cell lines: a comparative/collective analysis/study
Abstract
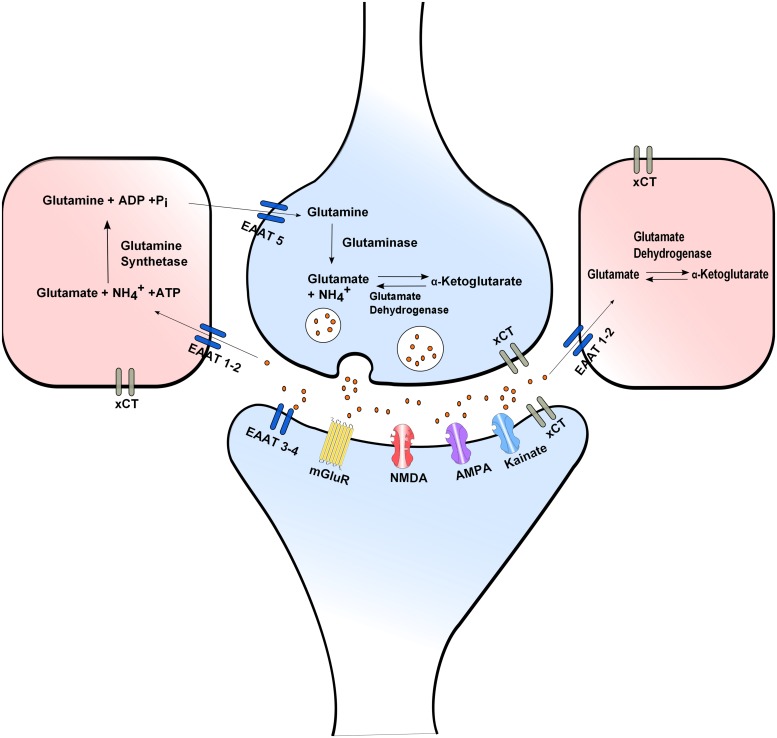
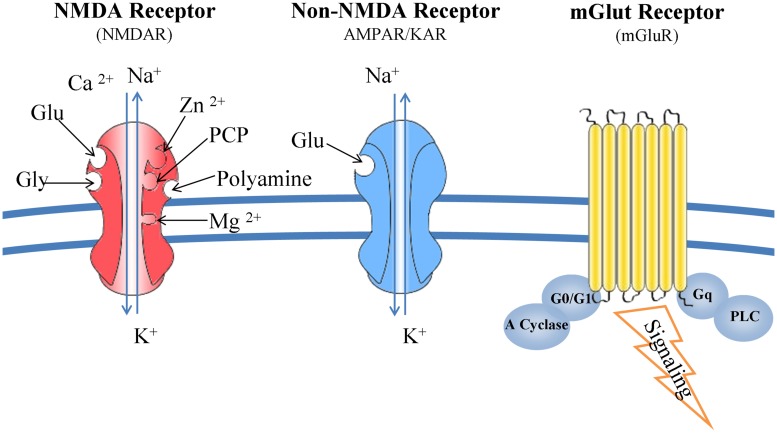
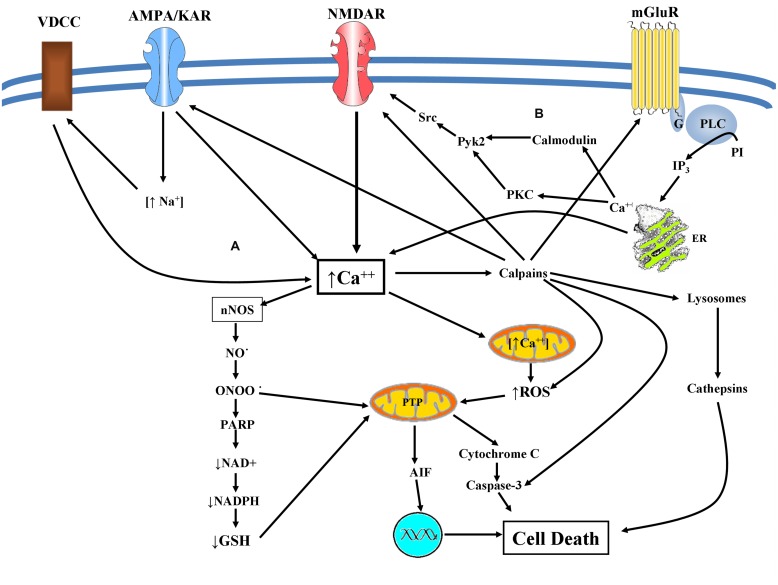
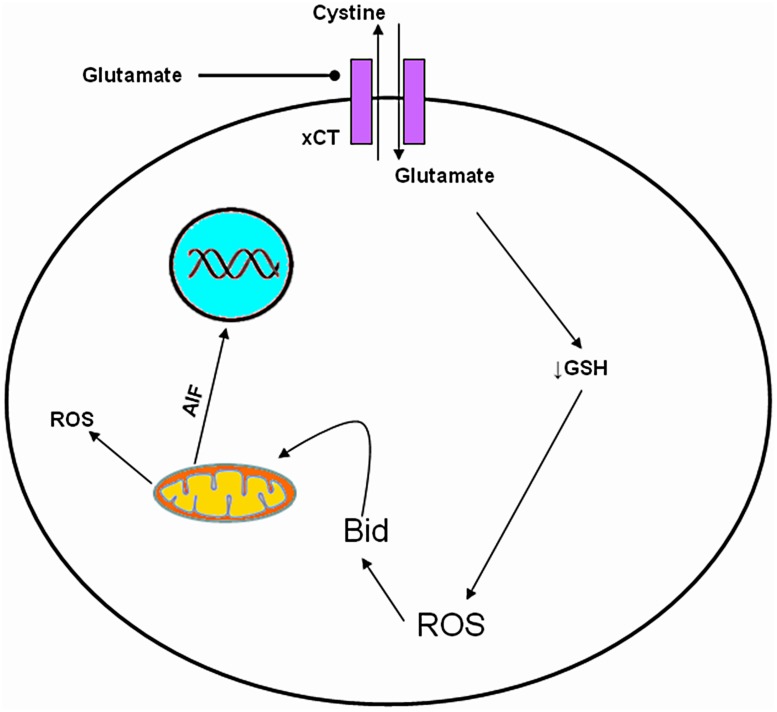
Although glutamate is one of the most important excitatory neurotransmitters of the central nervous system, its excessive extracellular concentration leads to uncontrolled continuous depolarization of neurons, a toxic process called, excitotoxicity. In excitotoxicity glutamate triggers the rise of intracellular Ca(2+) levels, followed by up regulation of nNOS, dysfunction of mitochondria, ROS production, ER stress, and release of lysosomal enzymes. Excessive calcium concentration is the key mediator of glutamate toxicity through over activation of ionotropic and metabotropic receptors. In addition, glutamate accumulation can also inhibit cystine (CySS) uptake by reversing the action of the CySS/glutamate antiporter. Reversal of the antiporter action reinforces the aforementioned events by depleting neurons of cysteine and eventually glutathione's reducing potential. Various cell lines have been employed in the pursuit to understand the mechanism(s) by which excitotoxicity affects the cells leading them ultimately to their demise. In some cell lines glutamate toxicity is exerted mainly through over activation of NMDA, AMPA, or kainate receptors whereas in other cell lines lacking such receptors, the toxicity is due to glutamate induced oxidative stress. However, in the greatest majority of the cell lines ionotropic glutamate receptors are present, co-existing to CySS/glutamate antiporters and metabotropic glutamate receptors, supporting the assumption that excitotoxicity effect in these cells is accumulative. Different cell lines differ in their responses when exposed to glutamate. In this review article the responses of PC12, SH-SY5Y, HT-22, NT-2, OLCs, C6, primary rat cortical neurons, RGC-5, and SCN2.2 cell systems are systematically collected and analyzed.
Keywords: HT-22; NT-2; PC12; RGC-5; SCN2.2; SH-SY5Y; excitotoxicity; glutamate oxidative toxicity.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous