

Dendritic patch-clamp recordings from cerebellar granule cells demonstrate electrotonic compactness
- PMID: 25852483
- PMCID: PMC4365719
- DOI: 10.3389/fncel.2015.00093
Dendritic patch-clamp recordings from cerebellar granule cells demonstrate electrotonic compactness
Abstract
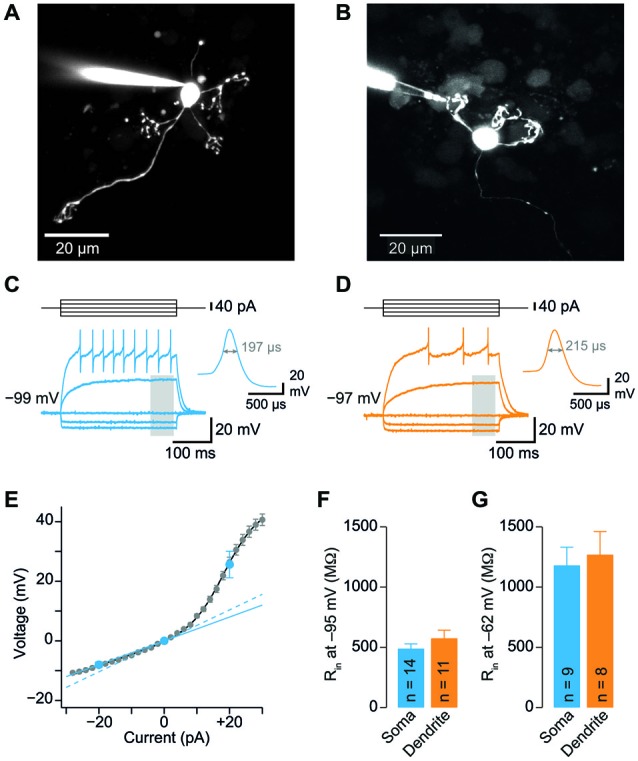
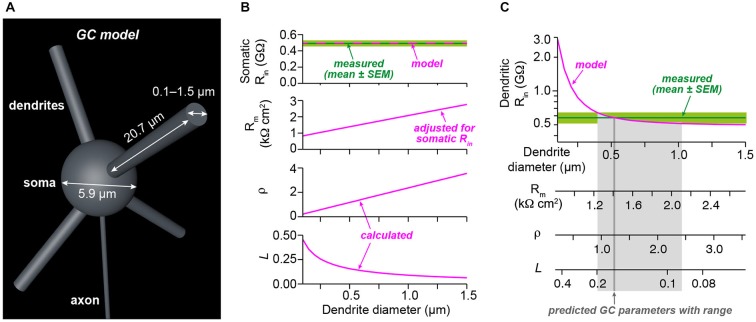
Cerebellar granule cells (GCs), the smallest neurons in the brain, have on average four short dendrites that receive high-frequency mossy fiber inputs conveying sensory information. The short length of the dendrites suggests that GCs are electrotonically compact allowing unfiltered integration of dendritic inputs. The small average diameter of the dendrites (~0.7 µm), however, argues for dendritic filtering. Previous studies based on somatic recordings and modeling indicated that GCs are electrotonically extremely compact. Here, we performed patch-clamp recordings from GC dendrites in acute brain slices of mice to directly analyze the electrotonic properties of GCs. Strikingly, the input resistance did not differ significantly between dendrites and somata of GCs. Furthermore, spontaneous excitatory postsynaptic potentials (EPSP) were similar in amplitude at dendritic and somatic recording sites. From the dendritic and somatic input resistances we determined parameters characterizing the electrotonic compactness of GCs. These data directly demonstrate that cerebellar GCs are electrotonically compact and thus ideally suited for efficient high-frequency information transfer.
Keywords: cerebellum; dendrites; electrophysiology; granule cell; patch-clamp techniques.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous