

Comparative whole-genome analysis of clinical isolates reveals characteristic architecture of Mycobacterium tuberculosis pangenome
- PMID: 25853708
- PMCID: PMC4390332
- DOI: 10.1371/journal.pone.0122979
Comparative whole-genome analysis of clinical isolates reveals characteristic architecture of Mycobacterium tuberculosis pangenome
Abstract
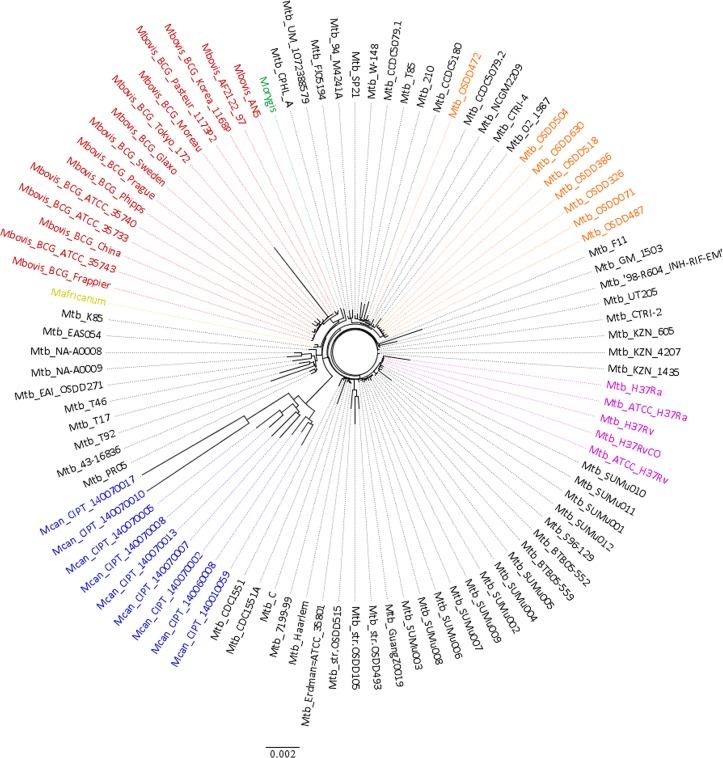
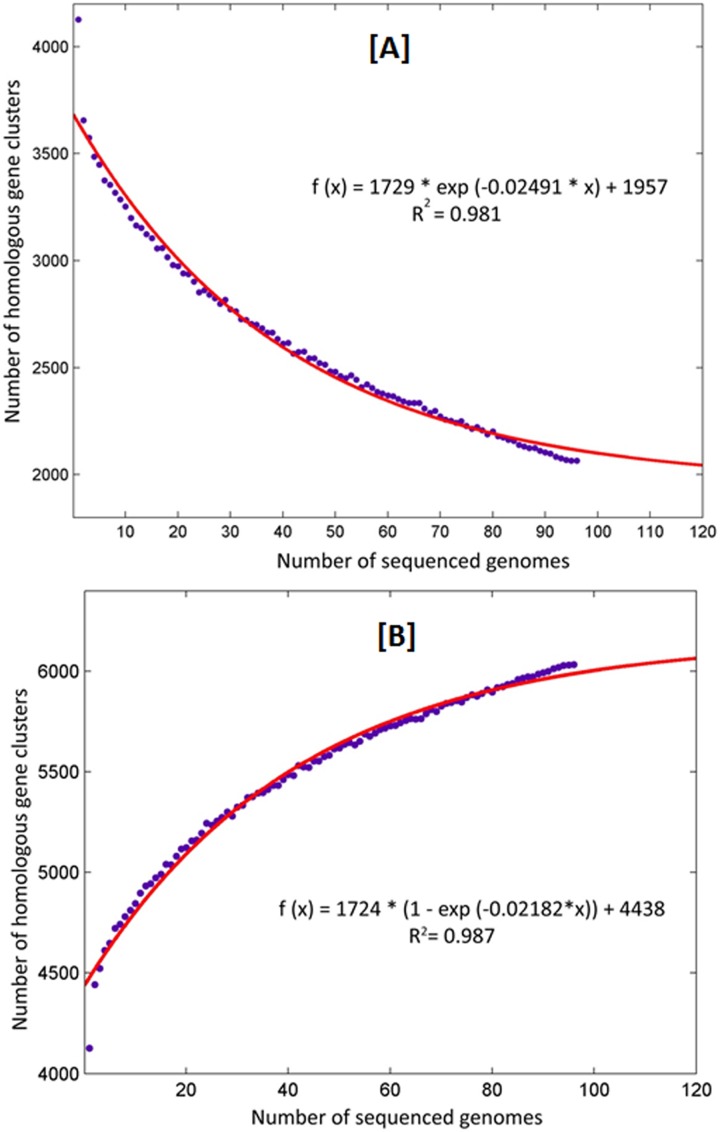
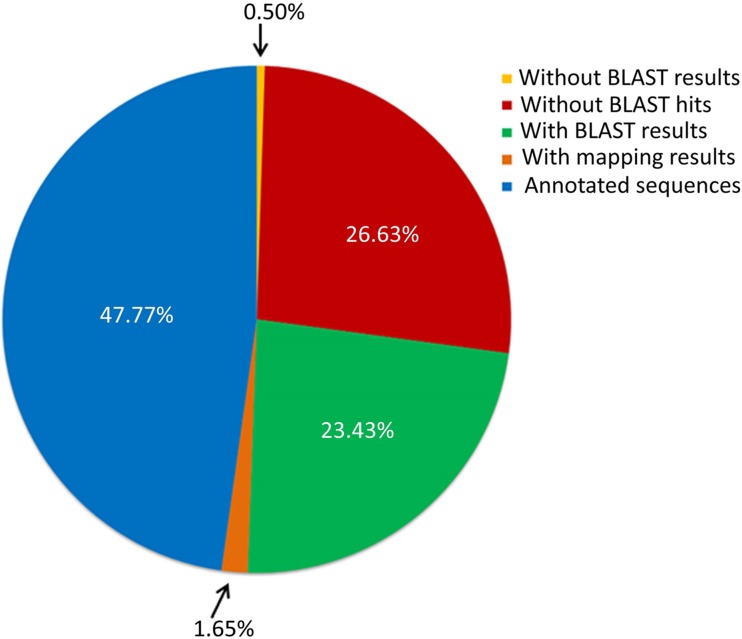
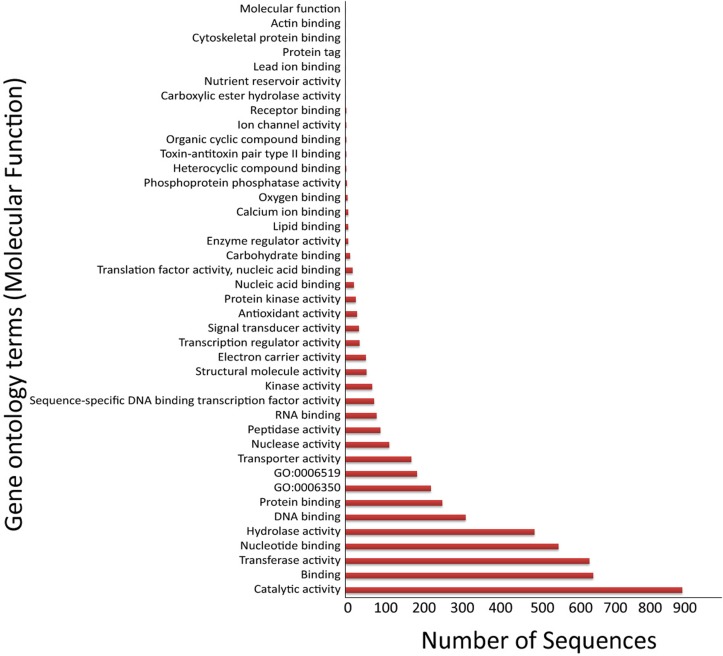
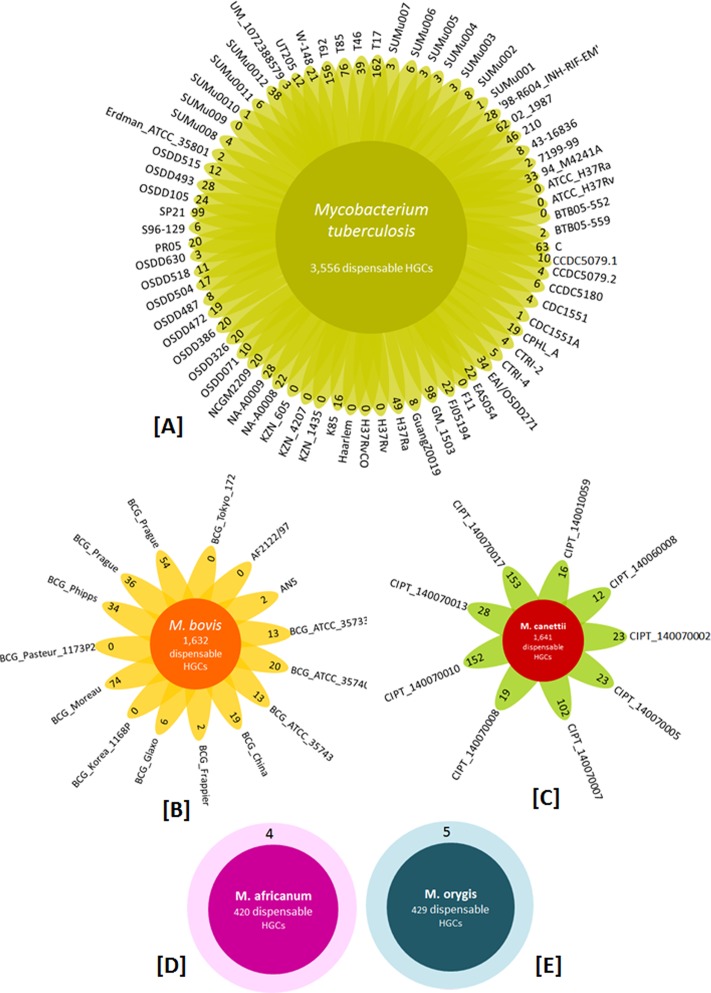
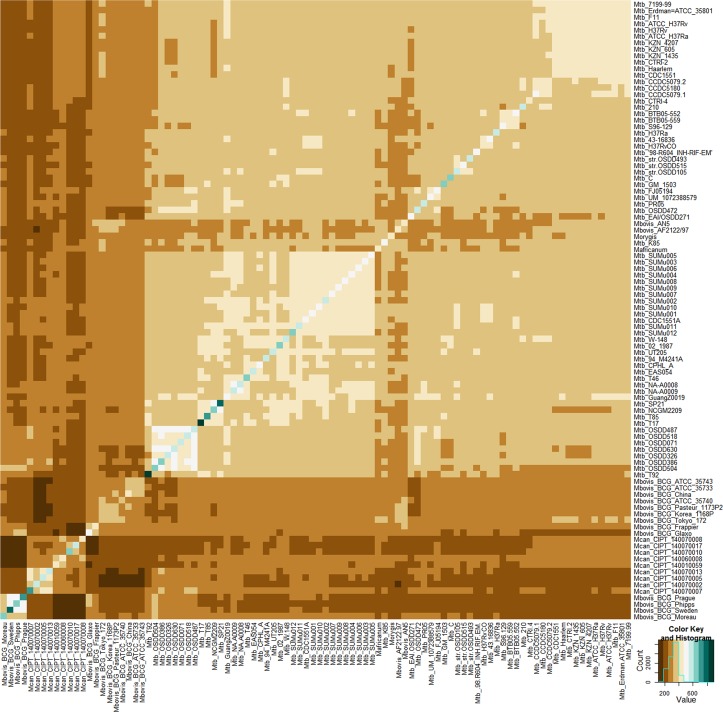
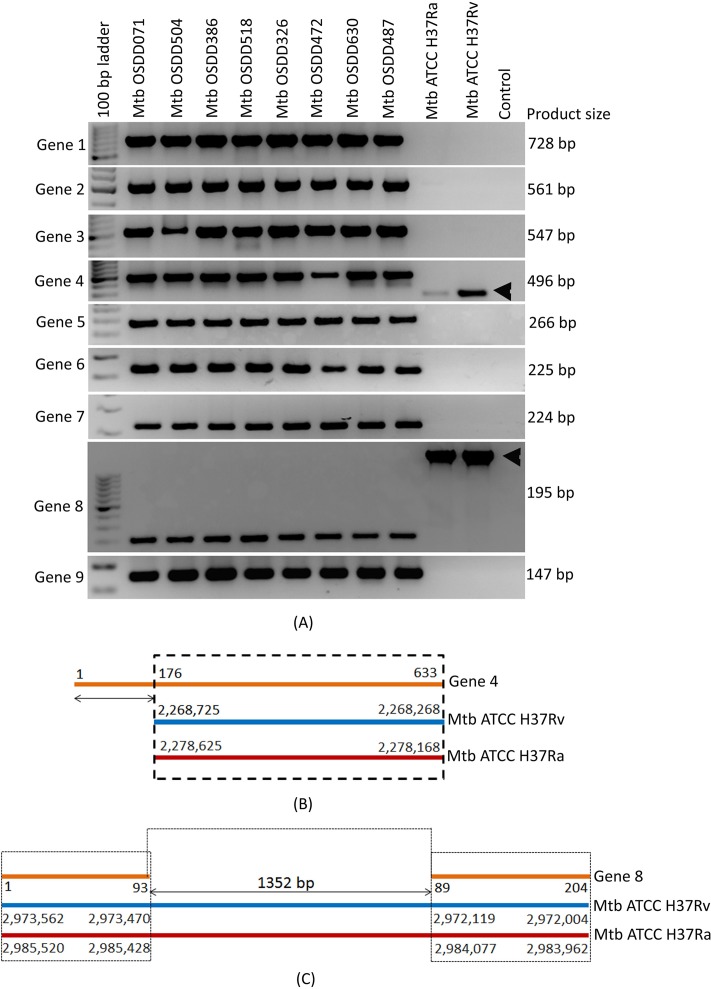
The tubercle complex consists of closely related mycobacterium species which appear to be variants of a single species. Comparative genome analysis of different strains could provide useful clues and insights into the genetic diversity of the species. We integrated genome assemblies of 96 strains from Mycobacterium tuberculosis complex (MTBC), which included 8 Indian clinical isolates sequenced and assembled in this study, to understand its pangenome architecture. We predicted genes for all the 96 strains and clustered their respective CDSs into homologous gene clusters (HGCs) to reveal a hard-core, soft-core and accessory genome component of MTBC. The hard-core (HGCs shared amongst 100% of the strains) was comprised of 2,066 gene clusters whereas the soft-core (HGCs shared amongst at least 95% of the strains) comprised of 3,374 gene clusters. The change in the core and accessory genome components when observed as a function of their size revealed that MTBC has an open pangenome. We identified 74 HGCs that were absent from reference strains H37Rv and H37Ra but were present in most of clinical isolates. We report PCR validation on 9 candidate genes depicting 7 genes completely absent from H37Rv and H37Ra whereas 2 genes shared partial homology with them accounting to probable insertion and deletion events. The pangenome approach is a promising tool for studying strain specific genetic differences occurring within species. We also suggest that since selecting appropriate target genes for typing purposes requires the expected target gene be present in all isolates being typed, therefore estimating the core-component of the species becomes a subject of prime importance.
Conflict of interest statement
Figures

Similar articles
-
Genetic Diversity of Mycobacterium tuberculosis Isolates from Assam, India: Dominance of Beijing Family and Discovery of Two New Clades Related to CAS1_Delhi and EAI Family Based on Spoligotyping and MIRU-VNTR Typing.PLoS One. 2015 Dec 23;10(12):e0145860. doi: 10.1371/journal.pone.0145860. eCollection 2015. PLoS One. 2015. PMID: 26701129 Free PMC article.
-
Identification of moaA3 gene in patient isolates of Mycobacterium tuberculosis in Kerala, which is absent in M. tuberculosis H37Rv and H37Ra.BMC Infect Dis. 2005 Oct 4;5:81. doi: 10.1186/1471-2334-5-81. BMC Infect Dis. 2005. PMID: 16202156 Free PMC article.
-
Mycobacterium tuberculosis complex lineage 5 exhibits high levels of within-lineage genomic diversity and differing gene content compared to the type strain H37Rv.Microb Genom. 2021 Jul;7(7):000437. doi: 10.1099/mgen.0.000437. Microb Genom. 2021. PMID: 34241588 Free PMC article.
-
Limitations of the Mycobacterium tuberculosis reference genome H37Rv in the detection of virulence-related loci.Genomics. 2017 Oct;109(5-6):471-474. doi: 10.1016/j.ygeno.2017.07.004. Epub 2017 Jul 22. Genomics. 2017. PMID: 28743540 Review.
-
Biological and Epidemiological Consequences of MTBC Diversity.Adv Exp Med Biol. 2017;1019:95-116. doi: 10.1007/978-3-319-64371-7_5. Adv Exp Med Biol. 2017. PMID: 29116631 Review.
Cited by
-
Decoding Ecuadorian Mycobacterium tuberculosis Isolates: Unveiling Lineage-Associated Signatures in Beta-Lactamase Resistance via Pangenome Analysis.Biomedicines. 2025 Jan 28;13(2):313. doi: 10.3390/biomedicines13020313. Biomedicines. 2025. PMID: 40002726 Free PMC article.
-
Mycobacterium bovis: From Genotyping to Genome Sequencing.Microorganisms. 2020 May 3;8(5):667. doi: 10.3390/microorganisms8050667. Microorganisms. 2020. PMID: 32375210 Free PMC article. Review.
-
Pan-genome association study of Mycobacterium tuberculosis lineage-4 revealed specific genes related to the high and low prevalence of the disease in patients from the North-Eastern area of Medellín, Colombia.Front Microbiol. 2023 Jan 4;13:1076797. doi: 10.3389/fmicb.2022.1076797. eCollection 2022. Front Microbiol. 2023. PMID: 36687645 Free PMC article.
-
A complete high-quality MinION nanopore assembly of an extensively drug-resistant Mycobacterium tuberculosis Beijing lineage strain identifies novel variation in repetitive PE/PPE gene regions.Microb Genom. 2018 Jul;4(7):e000188. doi: 10.1099/mgen.0.000188. Epub 2018 Jun 15. Microb Genom. 2018. PMID: 29906261 Free PMC article.
-
Pan and Core Genome Analysis of 183 Mycobacterium tuberculosis Strains Revealed a High Inter-Species Diversity among the Human Adapted Strains.Antibiotics (Basel). 2021 Apr 28;10(5):500. doi: 10.3390/antibiotics10050500. Antibiotics (Basel). 2021. PMID: 33924811 Free PMC article.
References
-
- de Jong BC, Antonio M, Gagneux S (2010) Mycobacterium africanum—review of an important cause of human tuberculosis in West Africa. PLoS Negl Trop Dis 4: e744 doi: 10.1371/journal.pntd.0000744 - DOI - PMC - PubMed
-
- Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, et al. (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393: 537–544. - PubMed
-
- van Ingen J, Rahim Z, Mulder A, Boeree MJ, Simeone R, Brosch R, et al. (2012) Characterization of Mycobacterium orygis as M. tuberculosis complex subspecies. Emerg Infect Dis 18: 653–655. doi: 610.3201/eid1804.110888 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
