

Plasticity of listeriolysin O pores and its regulation by pH and unique histidine [corrected]
- PMID: 25854672
- PMCID: PMC5381700
- DOI: 10.1038/srep09623
Plasticity of listeriolysin O pores and its regulation by pH and unique histidine [corrected]
Erratum in
-
Corrigendum: Plasticity of Listeriolysin O Pores and its Regulation by pH and Unique Histidine.Sci Rep. 2015 Nov 6;5:15690. doi: 10.1038/srep15690. Sci Rep. 2015. PMID: 26543014 Free PMC article. No abstract available.
Abstract
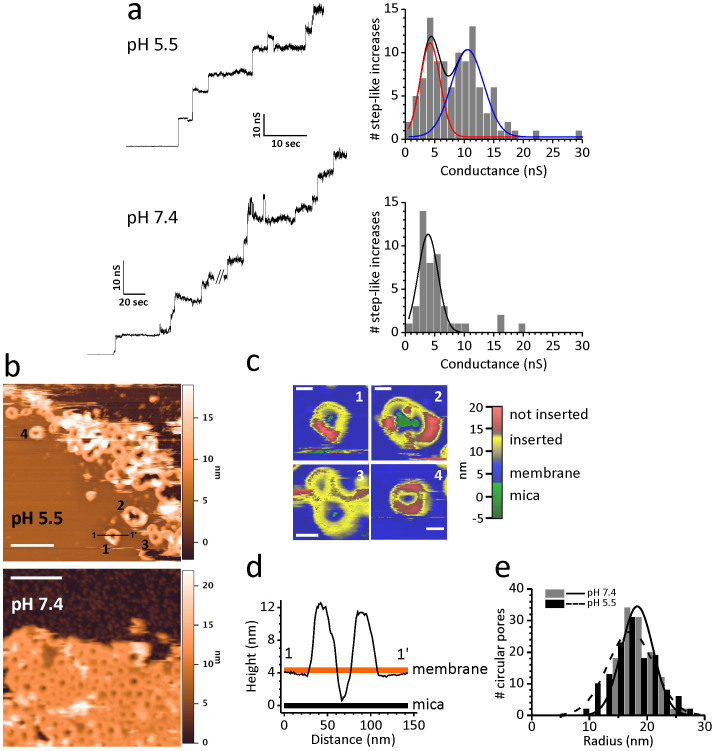
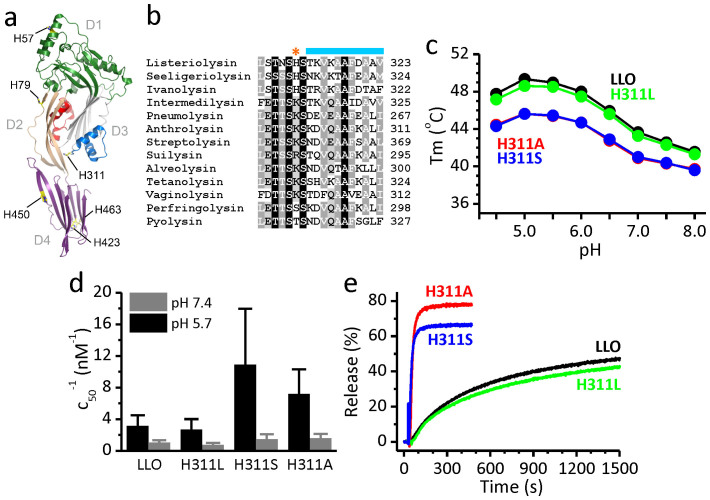
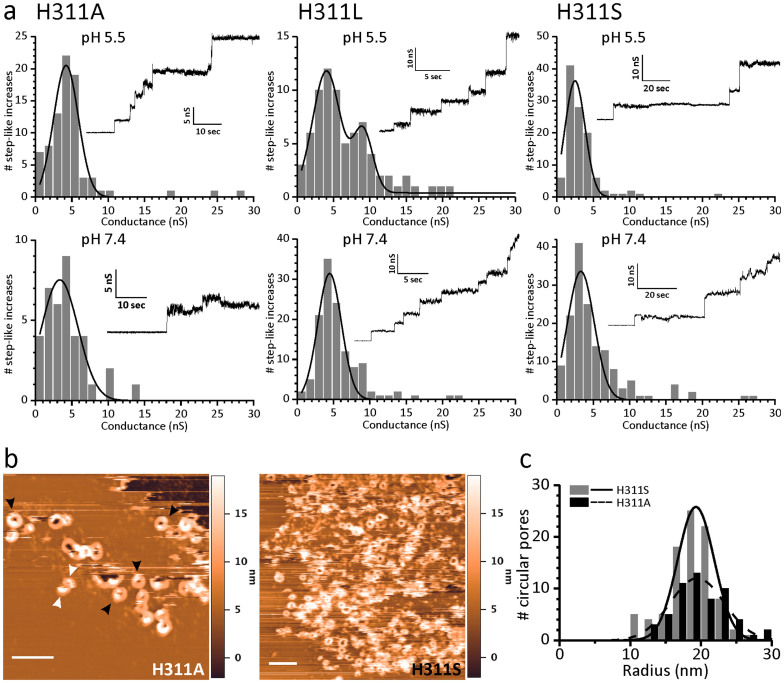
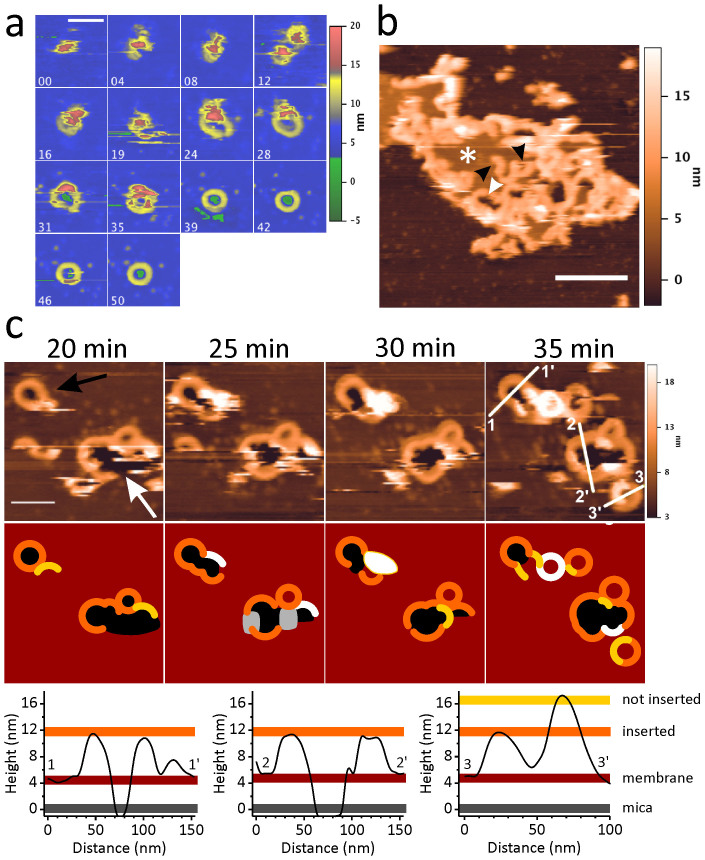
Pore formation of cellular membranes is an ancient mechanism of bacterial pathogenesis that allows efficient damaging of target cells. Several mechanisms have been described, however, relatively little is known about the assembly and properties of pores. Listeriolysin O (LLO) is a pH-regulated cholesterol-dependent cytolysin from the intracellular pathogen Listeria monocytogenes, which forms transmembrane β-barrel pores. Here we report that the assembly of LLO pores is rapid and efficient irrespective of pH. While pore diameters at the membrane surface are comparable at either pH 5.5 or 7.4, the distribution of pore conductances is significantly pH-dependent. This is directed by the unique residue H311, which is also important for the conformational stability of the LLO monomer and the rate of pore formation. The functional pores exhibit variations in height profiles and can reconfigure significantly by merging to other full pores or arcs. Our results indicate significant plasticity of large β-barrel pores, controlled by environmental cues like pH.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
