

Optogenetic evidence for inhibitory signaling from orexin to MCH neurons via local microcircuits
- PMID: 25855162
- PMCID: PMC4388912
- DOI: 10.1523/JNEUROSCI.5269-14.2015
Optogenetic evidence for inhibitory signaling from orexin to MCH neurons via local microcircuits
Abstract
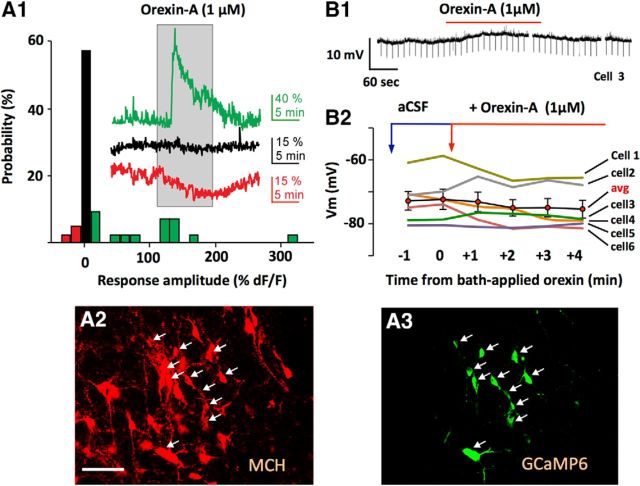
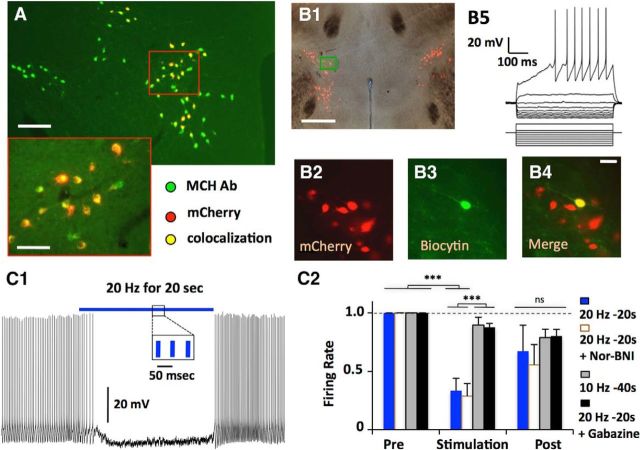
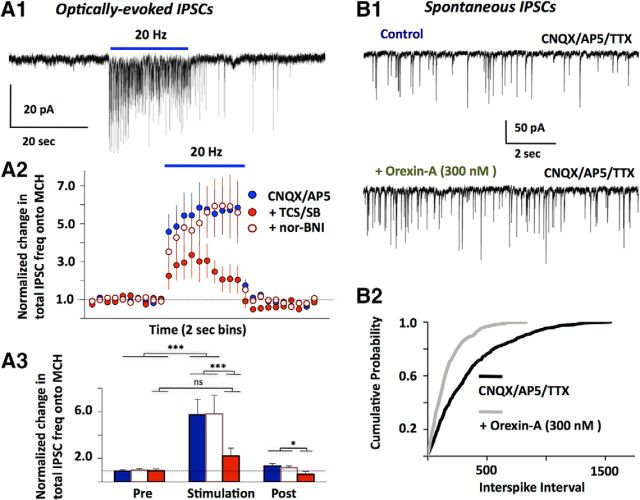
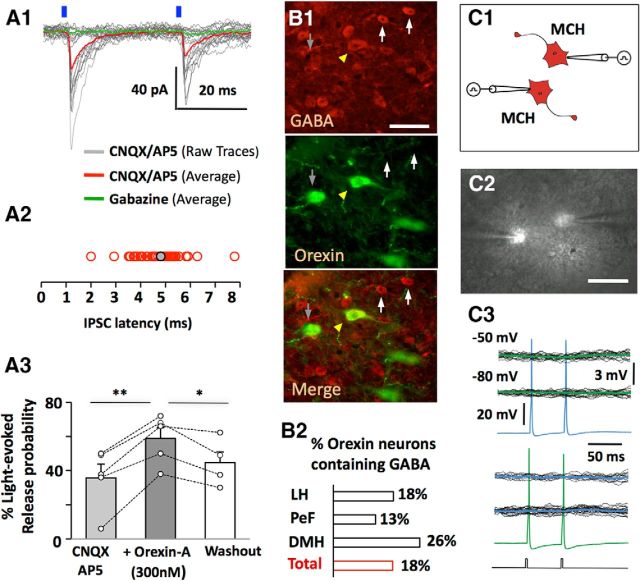
The lateral hypothalamus (LH) is a key regulator of multiple vital behaviors. The firing of brain-wide-projecting LH neurons releases neuropeptides promoting wakefulness (orexin/hypocretin; OH), or sleep (melanin-concentrating hormone; MCH). OH neurons, which coexpress glutamate and dynorphin, have been proposed to excite their neighbors, including MCH neurons, suggesting that LH may sometimes coengage its antagonistic outputs. However, it remains unclear if, when, and how OH actions promote temporal separation of the sleep and wake signals, a process that fails in narcolepsy caused by OH loss. To explore this directly, we paired optogenetic stimulation of OH cells (at rates that promoted awakening in vivo) with electrical monitoring of MCH cells in mouse brain slices. Membrane potential recordings showed that OH cell firing inhibited action potential firing in most MCH neurons, an effect that required GABAA but not dynorphin receptors. Membrane current analysis showed that OH cell firing increased the frequency of fast GABAergic currents in MCH cells, an effect blocked by antagonists of OH but not dynorphin or glutamate receptors, and mimicked by bath-applied OH peptide. In turn, neural network imaging with a calcium indicator genetically targeted to MCH neurons showed that excitation by bath-applied OH peptides occurs in a minority of MCH cells. Collectively, our data provide functional microcircuit evidence that intra-LH feedforward loops may facilitate appropriate switching between sleep and wake signals, potentially preventing sleep disorders.
Keywords: GABA; hypocretin; hypothalamus; melanin-concentrating hormone; optogenetics; orexin.
Copyright © 2015 the authors 0270-6474/15/355435-07$15.00/0.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources