

SHMT2 drives glioma cell survival in ischaemia but imposes a dependence on glycine clearance
- PMID: 25855294
- PMCID: PMC4533874
- DOI: 10.1038/nature14363
SHMT2 drives glioma cell survival in ischaemia but imposes a dependence on glycine clearance
Abstract
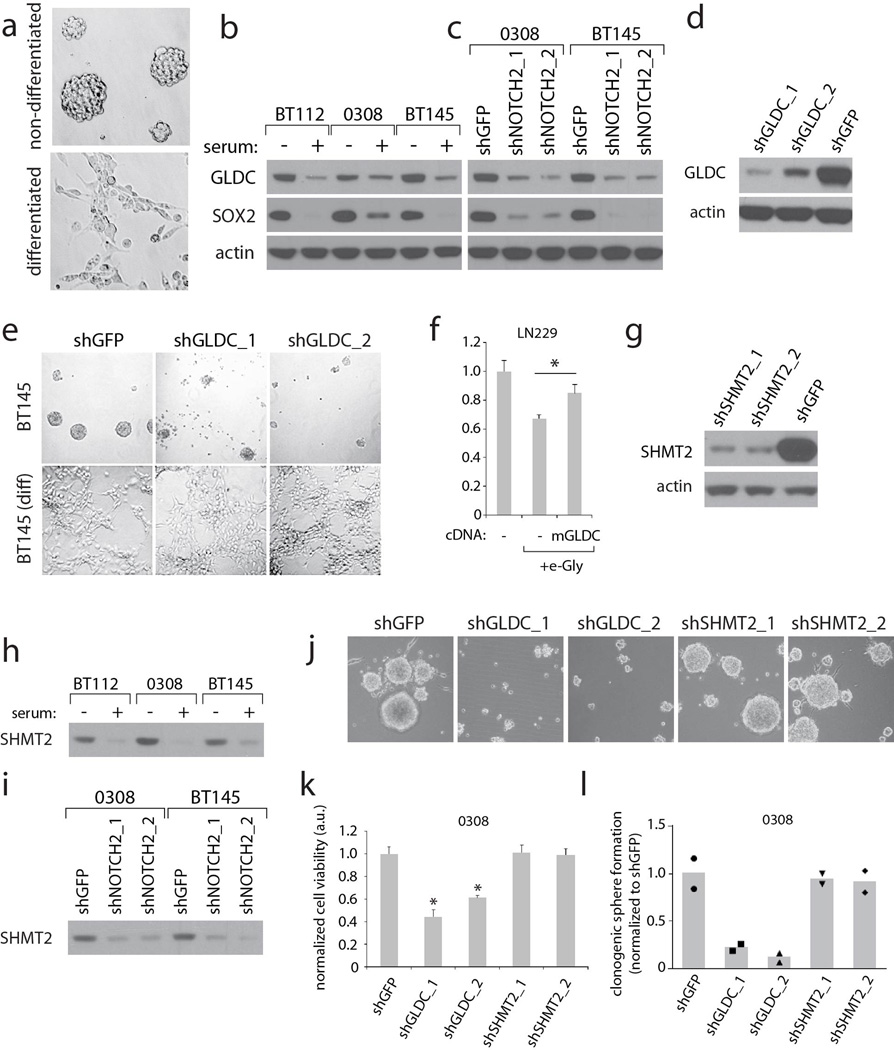
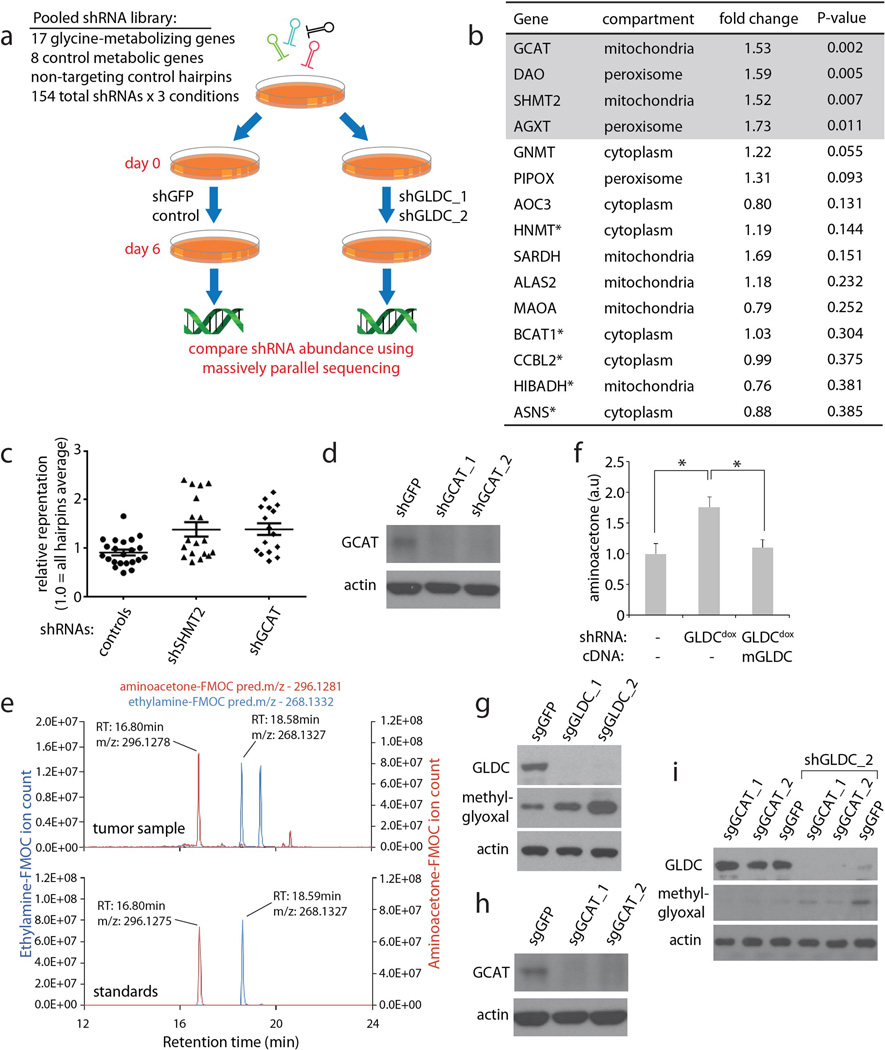
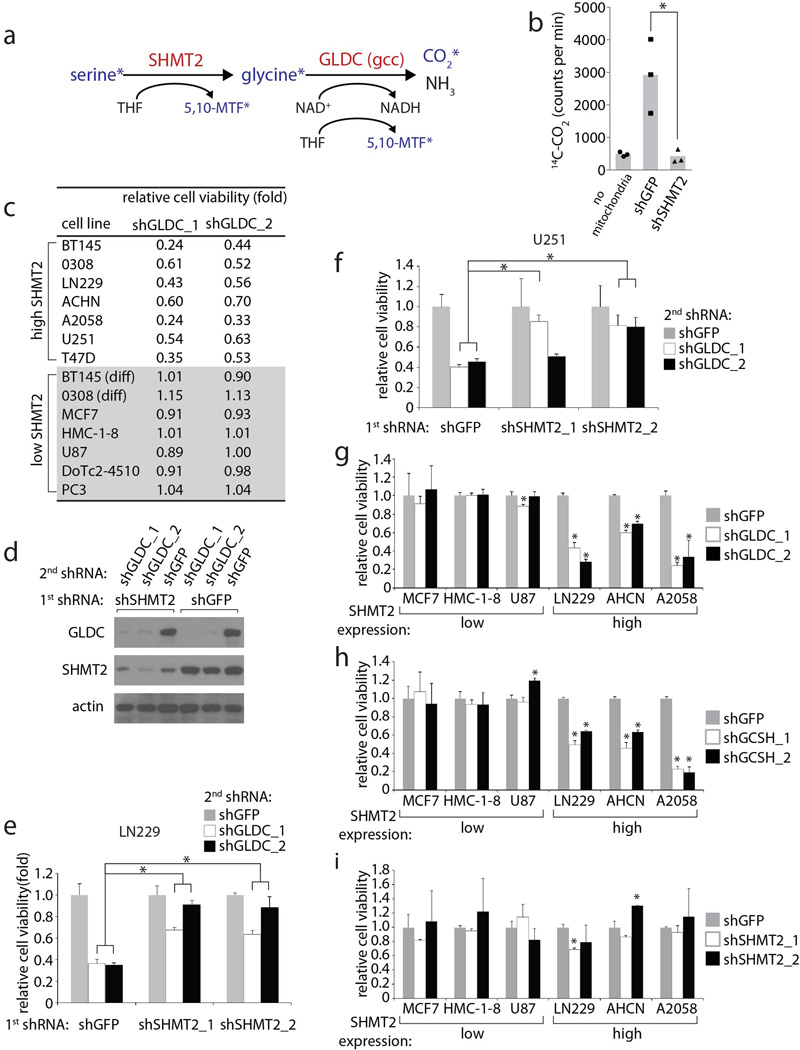
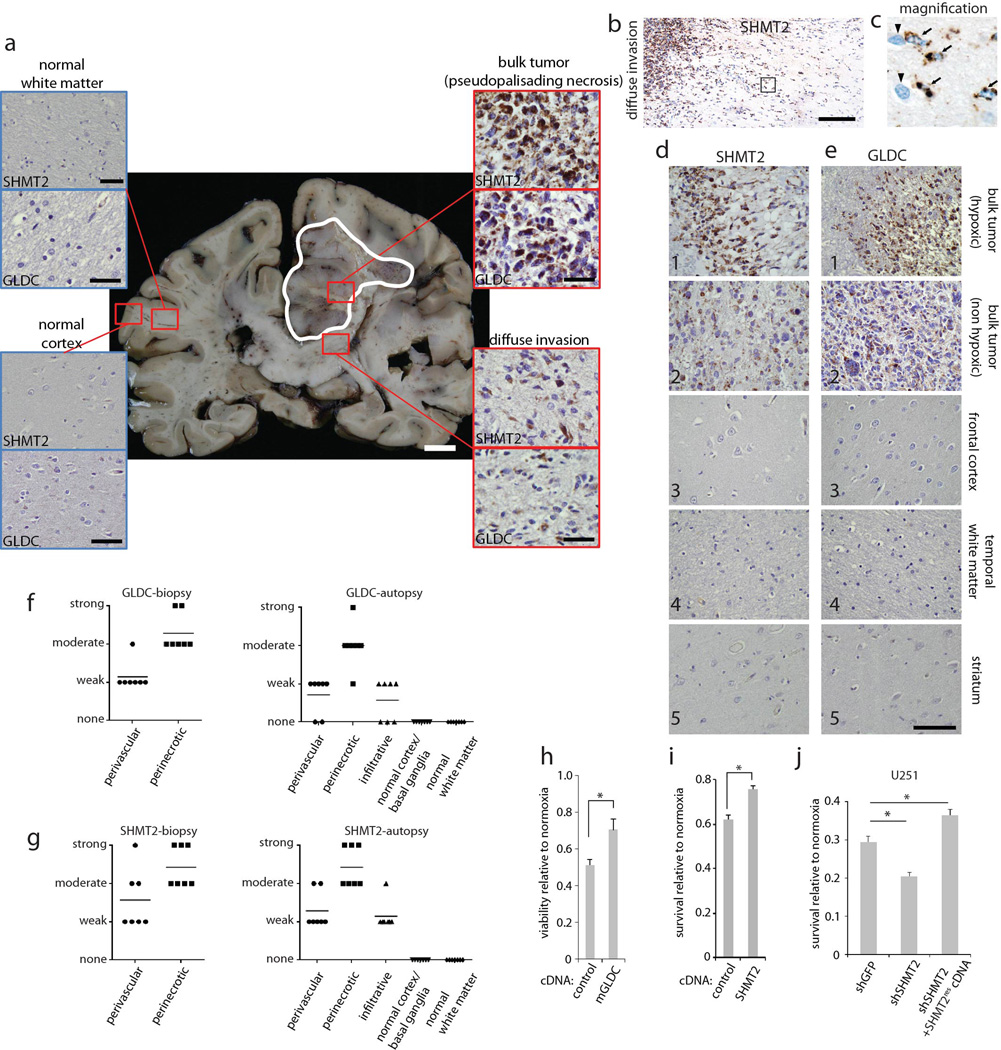
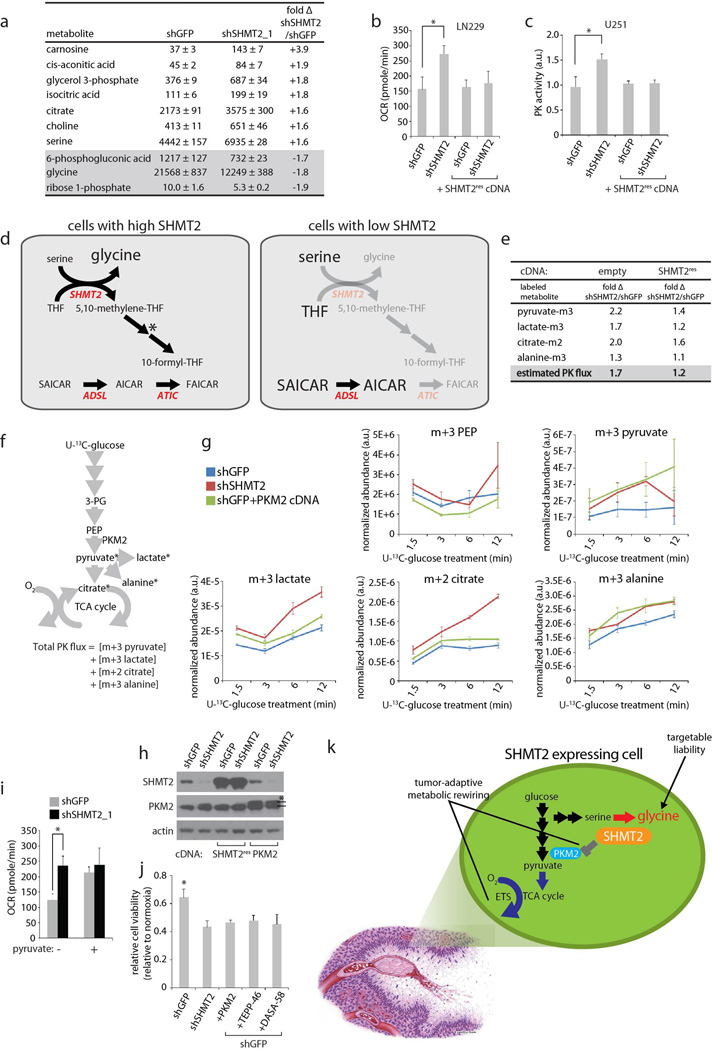
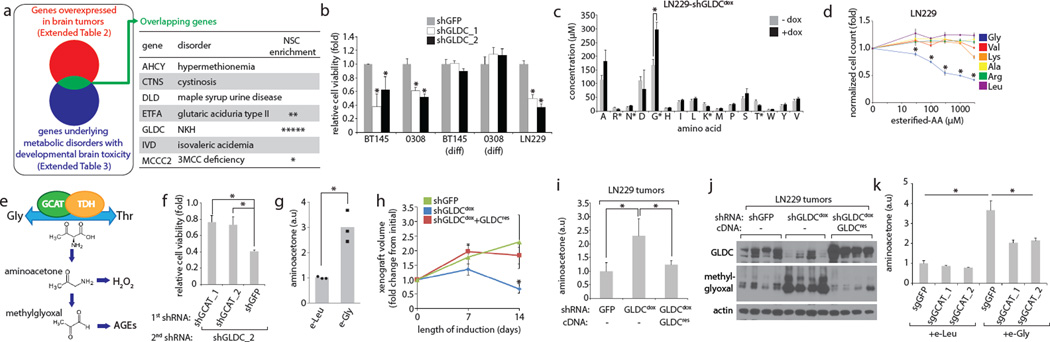
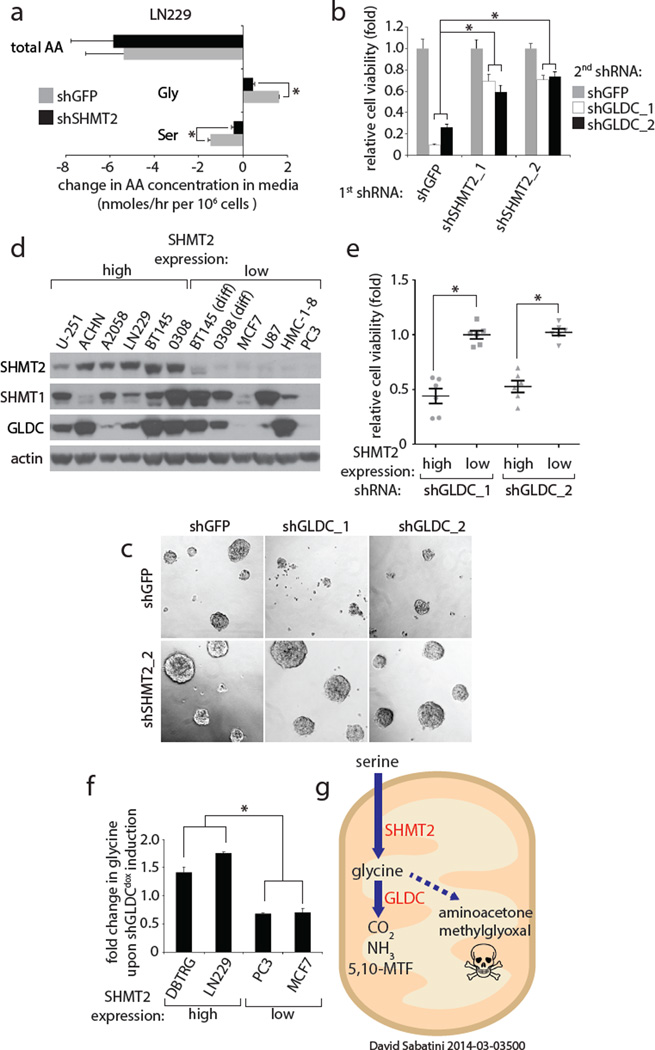
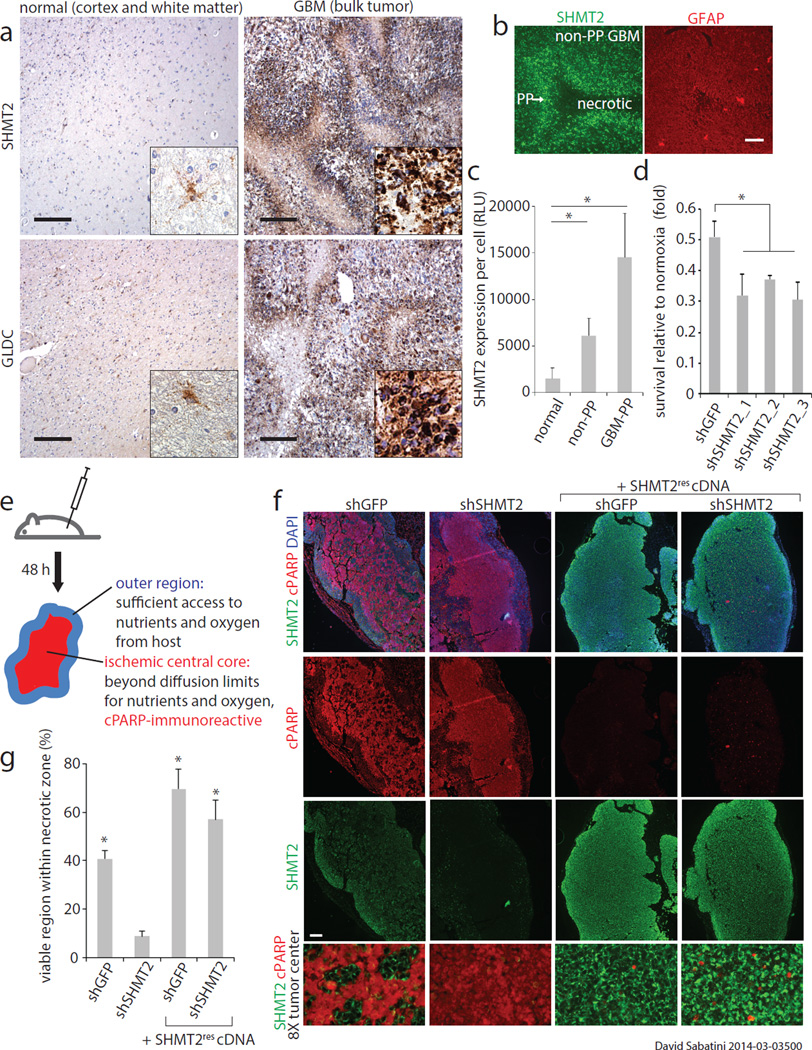
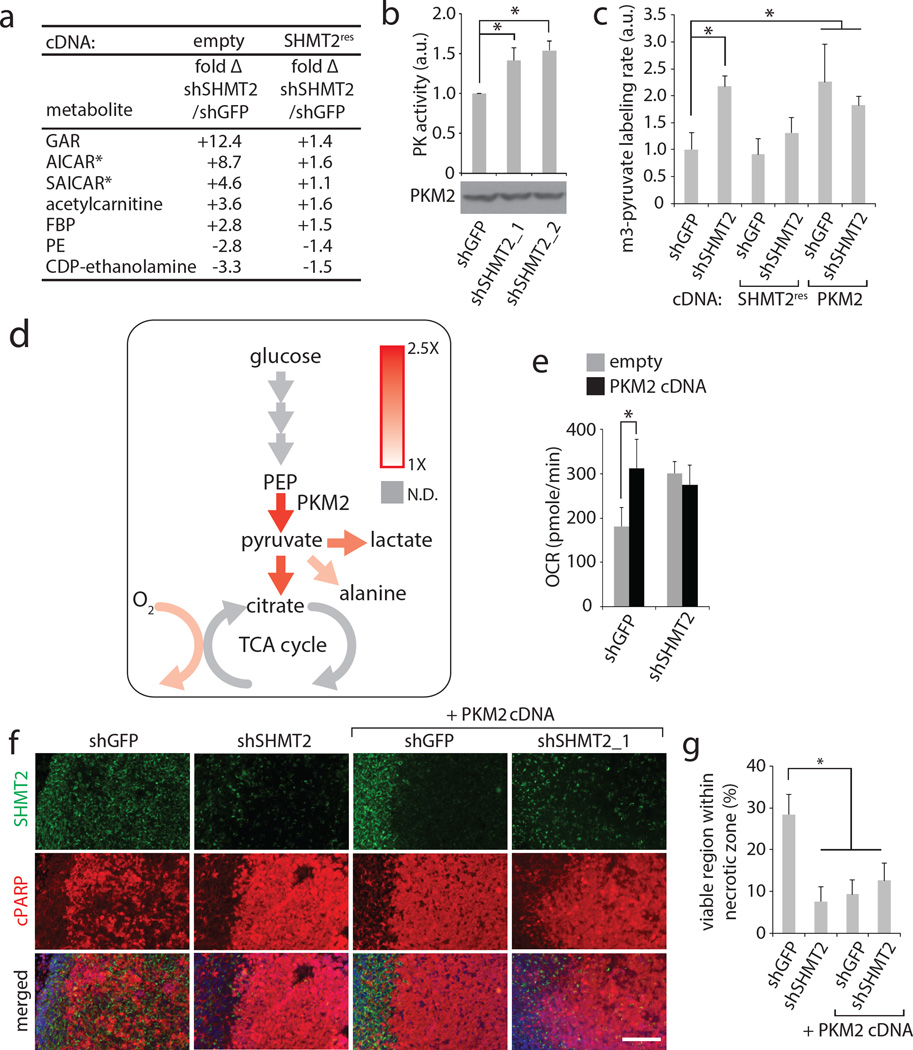
Cancer cells adapt their metabolic processes to support rapid proliferation, but less is known about how cancer cells alter metabolism to promote cell survival in a poorly vascularized tumour microenvironment. Here we identify a key role for serine and glycine metabolism in the survival of brain cancer cells within the ischaemic zones of gliomas. In human glioblastoma multiforme, mitochondrial serine hydroxymethyltransferase (SHMT2) and glycine decarboxylase (GLDC) are highly expressed in the pseudopalisading cells that surround necrotic foci. We find that SHMT2 activity limits that of pyruvate kinase (PKM2) and reduces oxygen consumption, eliciting a metabolic state that confers a profound survival advantage to cells in poorly vascularized tumour regions. GLDC inhibition impairs cells with high SHMT2 levels as the excess glycine not metabolized by GLDC can be converted to the toxic molecules aminoacetone and methylglyoxal. Thus, SHMT2 is required for cancer cells to adapt to the tumour environment, but also renders these cells sensitive to glycine cleavage system inhibition.
Figures
References
-
- Tennant DA, Duran RV, Gottlieb E. Targeting metabolic transformation for cancer therapy. Nat Rev Cancer. 2010;10:267–277. - PubMed
-
- Mattson MP, Shea TB. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003;26:137–146. doi:S0166-2236(03)00032-8 [pii] 10.1016/S0166-2236(03)00032-8. - PubMed
-
- Saudubray JM, Van den Berghe G, Walter J. Inborn metabolic diseases : diagnosis and treatment. 5th edn. Springer; 2012.
Supplementary References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA103866/CA/NCI NIH HHS/United States
- T32 GM007287/GM/NIGMS NIH HHS/United States
- P30 CA014051/CA/NCI NIH HHS/United States
- CA103866/CA/NCI NIH HHS/United States
- AI07389/AI/NIAID NIH HHS/United States
- R01CA168653/CA/NCI NIH HHS/United States
- 5P30CA14051/CA/NCI NIH HHS/United States
- T32GM007287/GM/NIGMS NIH HHS/United States
- R37 AI047389/AI/NIAID NIH HHS/United States
- R01 CA129105/CA/NCI NIH HHS/United States
- R00 CA168940/CA/NCI NIH HHS/United States
- T32 AI007389/AI/NIAID NIH HHS/United States
- K08-NS087118/NS/NINDS NIH HHS/United States
- R01 CA168653/CA/NCI NIH HHS/United States
- K99 CA168940/CA/NCI NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- K08 NS087118/NS/NINDS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- CA129105/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
