

Cellular level brain imaging in behaving mammals: an engineering approach
- PMID: 25856491
- PMCID: PMC5758309
- DOI: 10.1016/j.neuron.2015.03.055
Cellular level brain imaging in behaving mammals: an engineering approach
Abstract
Fluorescence imaging offers expanding capabilities for recording neural dynamics in behaving mammals, including the means to monitor hundreds of cells targeted by genetic type or connectivity, track cells over weeks, densely sample neurons within local microcircuits, study cells too inactive to isolate in extracellular electrical recordings, and visualize activity in dendrites, axons, or dendritic spines. We discuss recent progress and future directions for imaging in behaving mammals from a systems engineering perspective, which seeks holistic consideration of fluorescent indicators, optical instrumentation, and computational analyses. Today, genetically encoded indicators of neural Ca(2+) dynamics are widely used, and those of trans-membrane voltage are rapidly improving. Two complementary imaging paradigms involve conventional microscopes for studying head-restrained animals and head-mounted miniature microscopes for imaging in freely behaving animals. Overall, the field has attained sufficient sophistication that increased cooperation between those designing new indicators, light sources, microscopes, and computational analyses would greatly benefit future progress.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
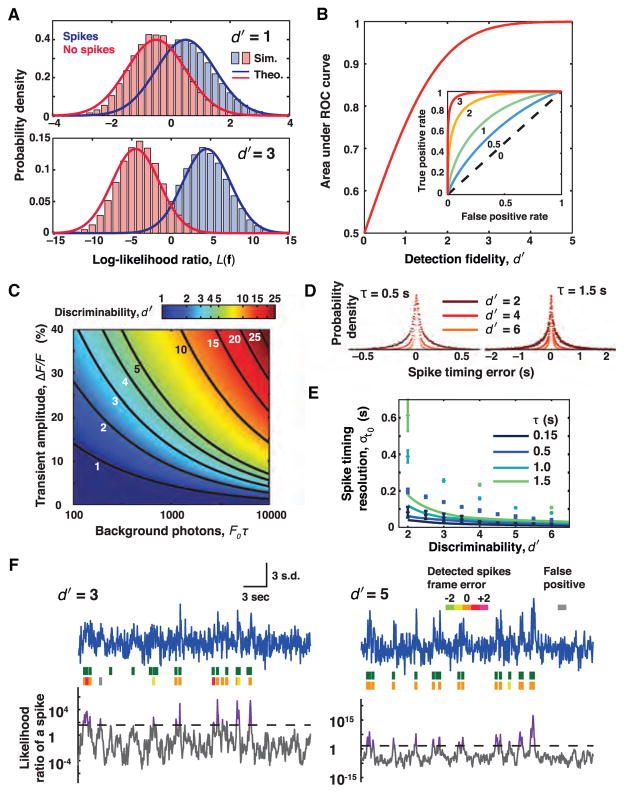
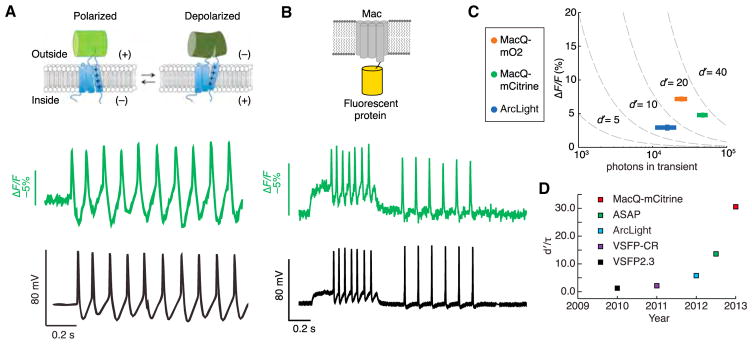

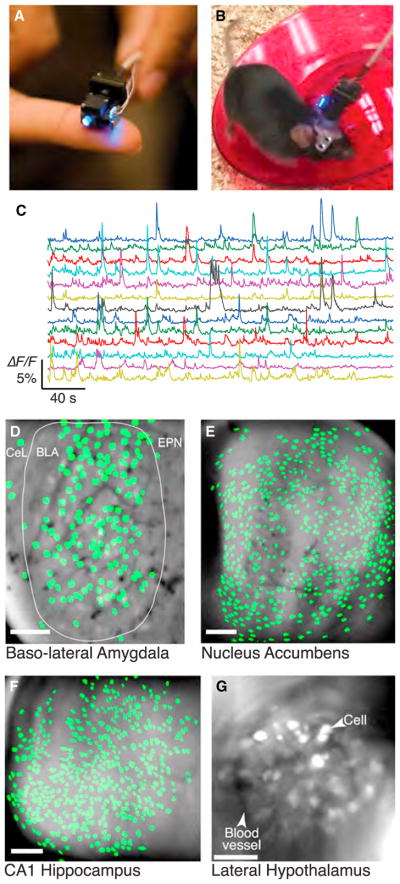

References
-
- Ahrens MB, Orger MB, Robson DN, Li JM, Keller PJ. Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat Methods. 2013;10:413–420. - PubMed
-
- Akemann W, Mutoh H, Perron A, Rossier J, Knöpfel T. Imaging brain electric signals with genetically targeted voltage-sensitive fluorescent proteins. Nat Methods. 2010;7:643–649. - PubMed
-
- Akemann W, Mutoh H, Perron A, Park YK, Iwamoto Y, Knöpfel T. Imaging neural circuit dynamics with a voltage-sensitive fluorescent protein. J Neurophysiol. 2012;108:2323–2337. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
