

The ventral pallidum: Subregion-specific functional anatomy and roles in motivated behaviors
- PMID: 25857550
- PMCID: PMC4687907
- DOI: 10.1016/j.pneurobio.2015.03.005
The ventral pallidum: Subregion-specific functional anatomy and roles in motivated behaviors
Abstract
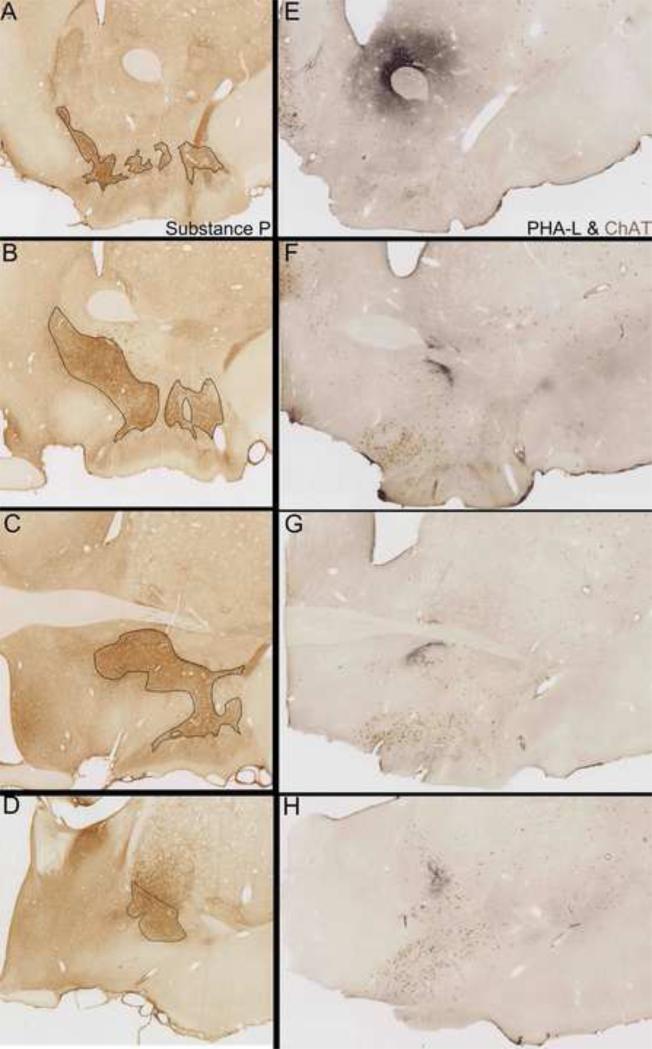
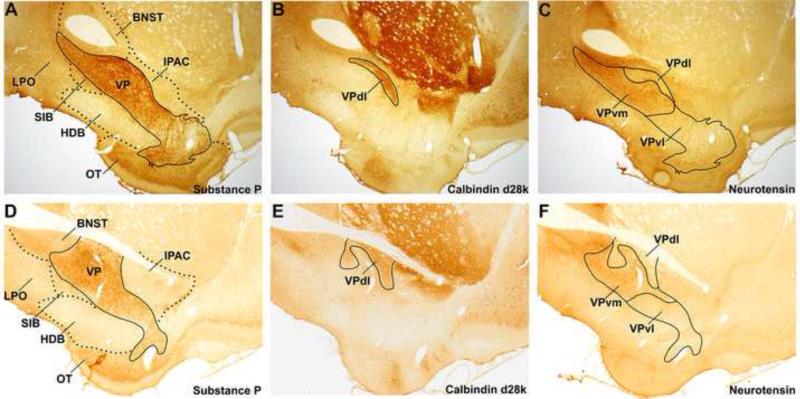
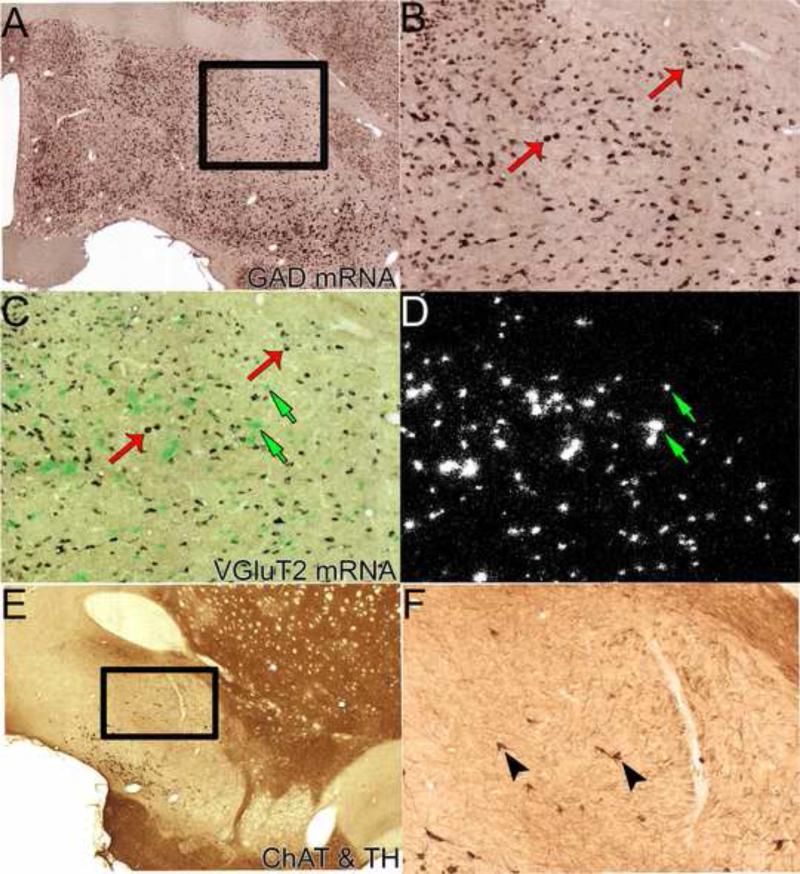
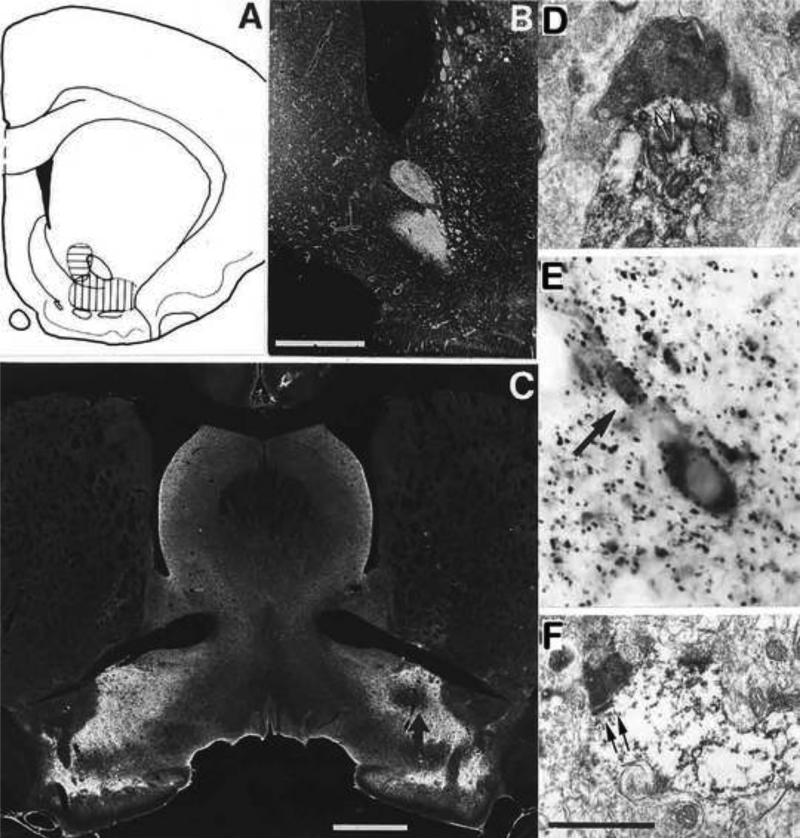
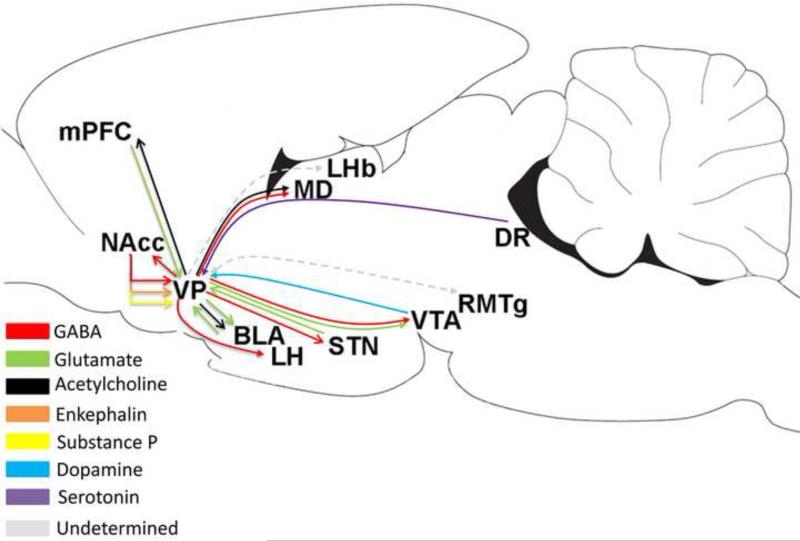
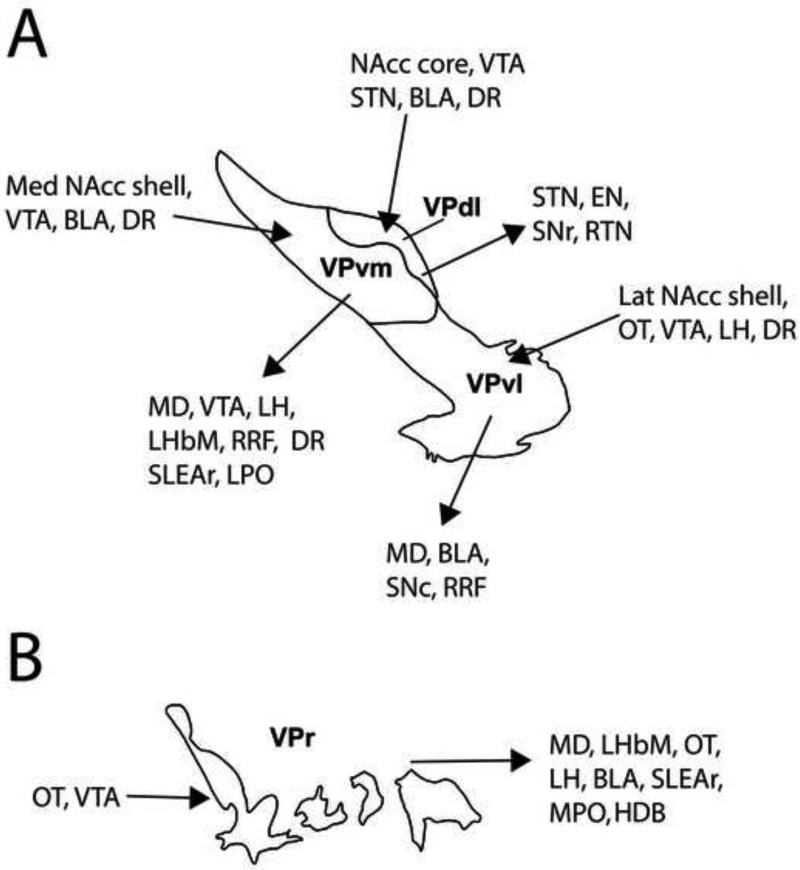
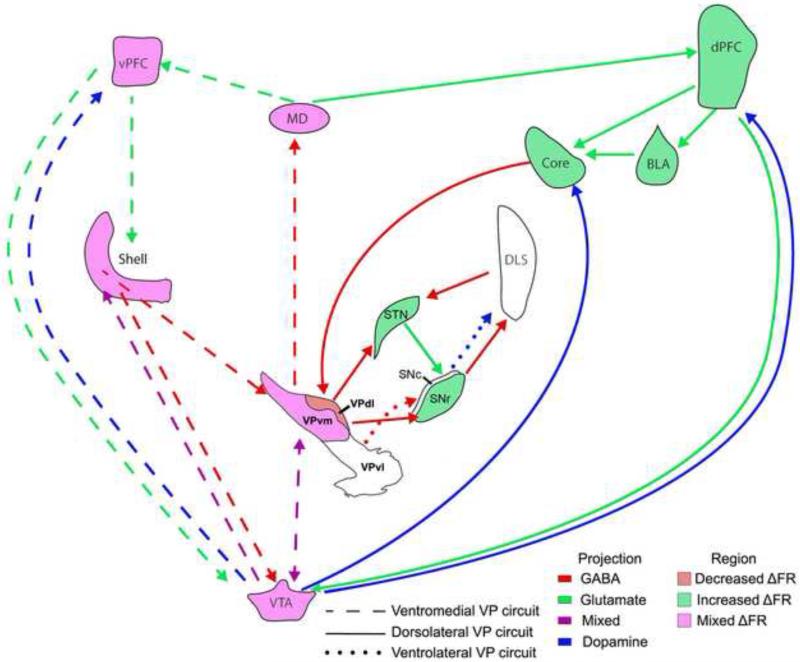
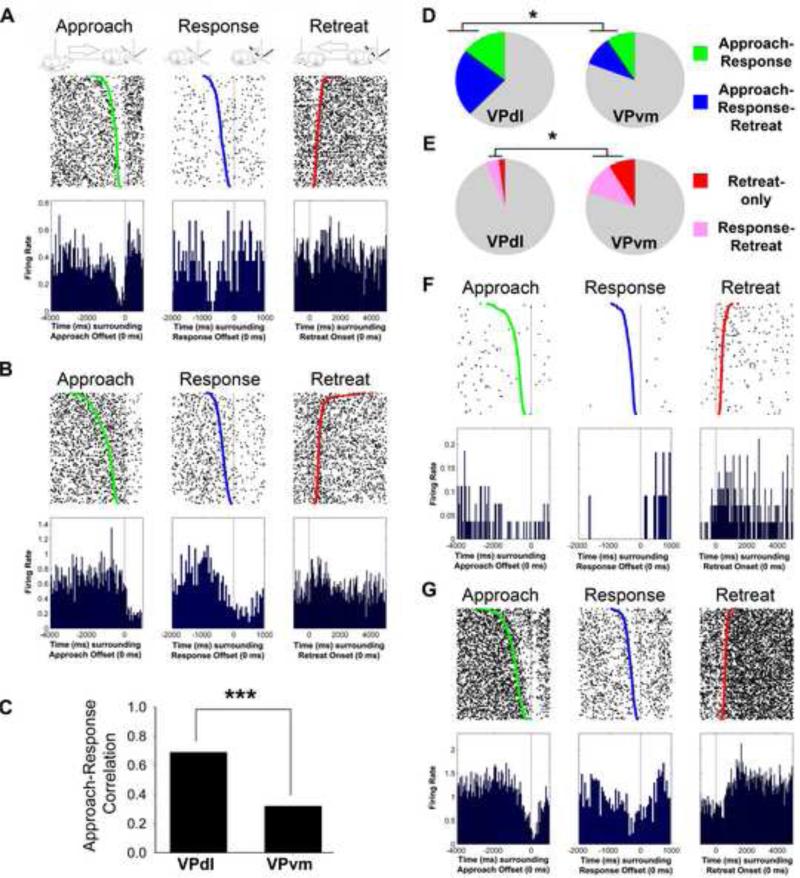
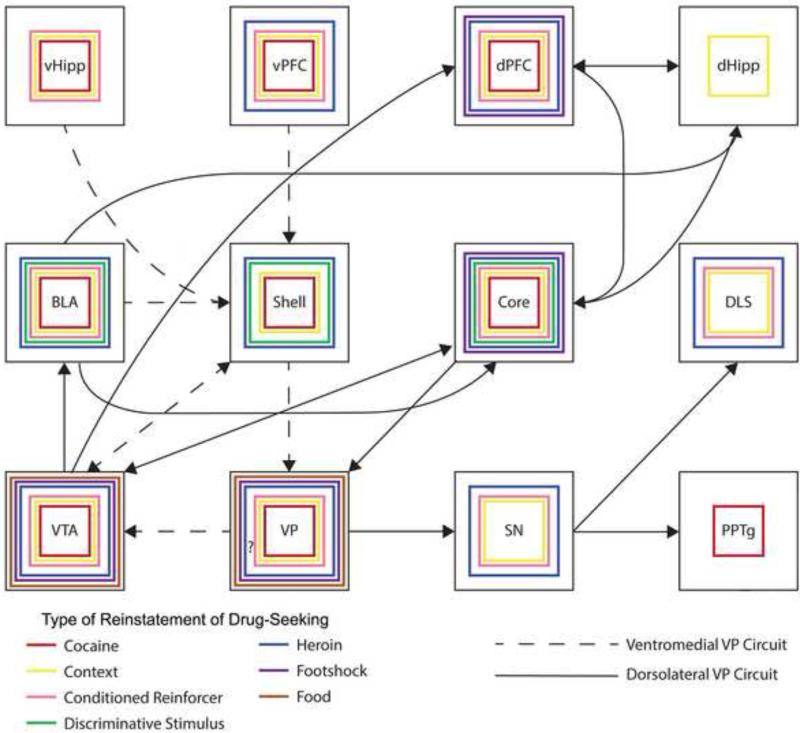
The ventral pallidum (VP) plays a critical role in the processing and execution of motivated behaviors. Yet this brain region is often overlooked in published discussions of the neurobiology of mental health (e.g., addiction, depression). This contributes to a gap in understanding the neurobiological mechanisms of psychiatric disorders. This review is presented to help bridge the gap by providing a resource for current knowledge of VP anatomy, projection patterns and subregional circuits, and how this organization relates to the function of VP neurons and ultimately behavior. For example, ventromedial (VPvm) and dorsolateral (VPdl) VP subregions receive projections from nucleus accumbens shell and core, respectively. Inhibitory GABAergic neurons of the VPvm project to mediodorsal thalamus, lateral hypothalamus, and ventral tegmental area, and this VP subregion helps discriminate the appropriate conditions to acquire natural rewards or drugs of abuse, consume preferred foods, and perform working memory tasks. GABAergic neurons of the VPdl project to subthalamic nucleus and substantia nigra pars reticulata, and this VP subregion is modulated by, and is necessary for, drug-seeking behavior. Additional circuits arise from nonGABAergic neuronal phenotypes that are likely to excite rather than inhibit their targets. These subregional and neuronal phenotypic circuits place the VP in a unique position to process motivationally relevant stimuli and coherent adaptive behaviors.
Keywords: Addiction; Dopamine; GABA; Glutamate; Nucleus accumbens; Ventral tegmental area.
Published by Elsevier Ltd.
Figures
References
-
- Alesdatter JE, Kalivas PW. Inhibition of mu opioid-induced motor activity in the ventral pallidum by D1 receptor blockade. Behav Pharmacol. 1993;4(6):645–651. - PubMed
-
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. - PubMed
-
- Alheid GF. Extended amygdala and basal forebrain. Ann NY Acad Sci. 2003;985:185–205. - PubMed
-
- Alheid GF, Heimer L. New perspectives in basal forebrain organization of special relevance for neuropsychiatric disorders: the striatopallidal, amygdaloid, and corticopetal components of substantia innominata. Neuroscience. 1988;27(1):1–39. - PubMed
-
- Alleweireldt AT, Hobbs RJ, Taylor AR, Neisewander JL. Effects of SCH-23390 infused into the amygdala or adjacent cortex and basal ganglia on cocaine seeking and self-administration in rats. Neuropsychopharmacology. 2006;31(2):363–74. - PubMed
Publication types
MeSH terms
Grants and funding
- R01 DA015760/DA/NIDA NIH HHS/United States
- F31 DA023306/DA/NIDA NIH HHS/United States
- DA021475/DA/NIDA NIH HHS/United States
- R01 DA005255/DA/NIDA NIH HHS/United States
- NS23945/NS/NINDS NIH HHS/United States
- DA05255/DA/NIDA NIH HHS/United States
- DA019783/DA/NIDA NIH HHS/United States
- R01 DA006886/DA/NIDA NIH HHS/United States
- F31 DA019763/DA/NIDA NIH HHS/United States
- F31 DA021475/DA/NIDA NIH HHS/United States
- DA023306/DA/NIDA NIH HHS/United States
- F32 DA005651/DA/NIDA NIH HHS/United States
- F32 DA05651/DA/NIDA NIH HHS/United States
- F31 DA019783/DA/NIDA NIH HHS/United States
- DA015760/DA/NIDA NIH HHS/United States
- DA024923/DA/NIDA NIH HHS/United States
- F31 DA026252/DA/NIDA NIH HHS/United States
- R01 NS023945/NS/NINDS NIH HHS/United States
- DA026252/DA/NIDA NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- DA0331231/DA/NIDA NIH HHS/United States
- F31 DA024923/DA/NIDA NIH HHS/United States
- DA006886/DA/NIDA NIH HHS/United States
- RF1 NS023945/NS/NINDS NIH HHS/United States
- F30 MH45180/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
