

TNF-related apoptosis-inducing ligand promotes human preadipocyte proliferation via ERK1/2 activation
- PMID: 25857555
- PMCID: PMC4478800
- DOI: 10.1096/fj.14-267278
TNF-related apoptosis-inducing ligand promotes human preadipocyte proliferation via ERK1/2 activation
Abstract
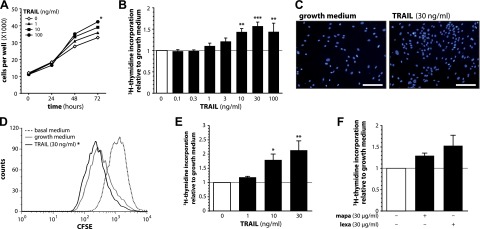
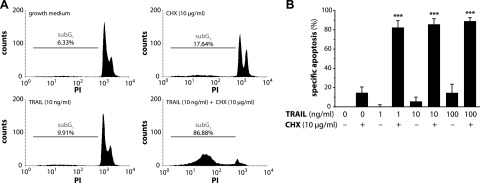
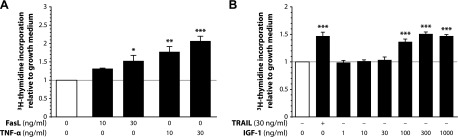
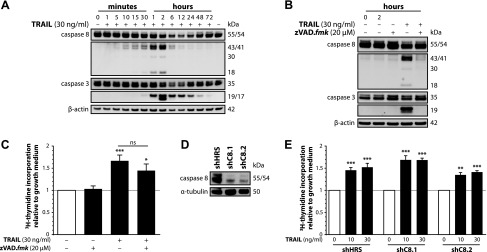
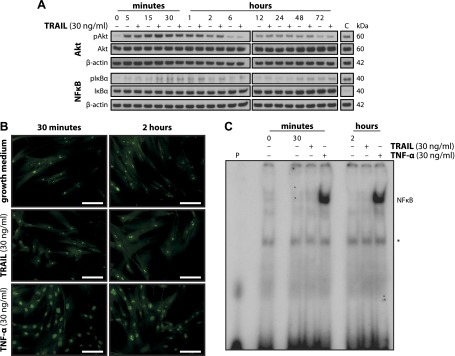
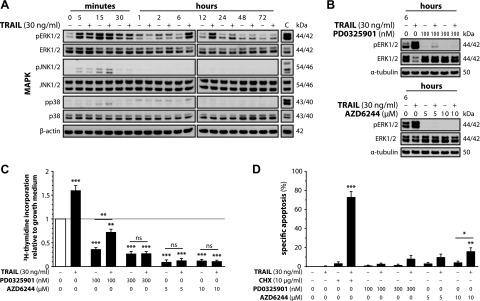
Upon obesity, adipose tissue is excessively expanded and characterized by pathologic processes like hypoxia, fibrosis, and inflammation. Death ligands belonging to the TNF superfamily such as TNF-α are important contributors to these derangements and exert a pronounced influence on the metabolic and cellular homeostasis of adipose tissue. Here, we sought to identify the effect of the death ligand TNF-related apoptosis-inducing ligand (TRAIL) on the adipose tissue precursor cell pool and therefore investigated its influence on preadipocyte proliferation. Treatment of human preadipocytes with TRAIL resulted in a time- and dose-dependent increase in proliferation (EC50 3.4 ng/ml) comparable to IGF-1. Although no apoptosis was observed, TRAIL triggered a rapid cleavage of caspase-8 and -3. Neither inhibition of caspase activity by zVAD.fmk (20 µM) nor ablation of caspase-8 expression by lentivirus-delivered small hairpin RNA (shRNA) abolished the proliferative response. TRAIL triggered a delayed and sustained activation of ERK1/2, leaving Akt, p38, JNK, and NF-κB unaffected. Importantly, inhibition of ERK1/2 activation by PD0325901 (300 nM) or AZD6244 (5 or 10 µM) completely abolished the proliferative response. We thus reveal a hitherto unknown function of TRAIL in regulating adipose tissue homeostasis by promoting the proliferation of tissue-resident precursor cells.
Keywords: adipocyte progenitor; adipose tissue homeostasis; death ligand; noncanonical signaling.
© FASEB.
Figures
Similar articles
-
TRAIL (TNF-related apoptosis-inducing ligand) inhibits human adipocyte differentiation via caspase-mediated downregulation of adipogenic transcription factors.Cell Death Dis. 2016 Oct 13;7(10):e2412. doi: 10.1038/cddis.2016.286. Cell Death Dis. 2016. PMID: 27735943 Free PMC article.
-
miR-146a-mediated suppression of the inflammatory response in human adipocytes.Sci Rep. 2016 Dec 6;6:38339. doi: 10.1038/srep38339. Sci Rep. 2016. PMID: 27922090 Free PMC article.
-
TRAIL (TNF-related apoptosis-inducing ligand) regulates adipocyte metabolism by caspase-mediated cleavage of PPARgamma.Cell Death Dis. 2013 Jan 24;4(1):e474. doi: 10.1038/cddis.2012.212. Cell Death Dis. 2013. PMID: 23348588 Free PMC article.
-
Trail (TNF-related apoptosis-inducing ligand) induces an inflammatory response in human adipocytes.Sci Rep. 2017 Jul 18;7(1):5691. doi: 10.1038/s41598-017-05932-7. Sci Rep. 2017. PMID: 28720906 Free PMC article.
-
MAPK p38 and JNK have opposing activities on TRAIL-induced apoptosis activation in NSCLC H460 cells that involves RIP1 and caspase-8 and is mediated by Mcl-1.Apoptosis. 2013 Jul;18(7):851-60. doi: 10.1007/s10495-013-0829-3. Apoptosis. 2013. PMID: 23456625
Cited by
-
A TRAIL-TL1A Paracrine Network Involving Adipocytes, Macrophages, and Lymphocytes Induces Adipose Tissue Dysfunction Downstream of E2F1 in Human Obesity.Diabetes. 2020 Nov;69(11):2310-2323. doi: 10.2337/db19-1231. Epub 2020 Jul 30. Diabetes. 2020. PMID: 32732304 Free PMC article.
-
Mechanisms Involved in Childhood Obesity-Related Bone Fragility.Front Endocrinol (Lausanne). 2019 May 3;10:269. doi: 10.3389/fendo.2019.00269. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31130918 Free PMC article. Review.
-
Inflammo-immune perspective on the association of eight migraine risk factors with migraine: a multi-omics Mendelian randomization study.Front Neurol. 2024 Aug 7;15:1440995. doi: 10.3389/fneur.2024.1440995. eCollection 2024. Front Neurol. 2024. PMID: 39170074 Free PMC article.
-
Beyond adiponectin and leptin: adipose tissue-derived mediators of inter-organ communication.J Lipid Res. 2019 Oct;60(10):1648-1684. doi: 10.1194/jlr.R094060. Epub 2019 Jun 17. J Lipid Res. 2019. PMID: 31209153 Free PMC article. Review.
-
TRAIL (TNF-related apoptosis-inducing ligand) inhibits human adipocyte differentiation via caspase-mediated downregulation of adipogenic transcription factors.Cell Death Dis. 2016 Oct 13;7(10):e2412. doi: 10.1038/cddis.2016.286. Cell Death Dis. 2016. PMID: 27735943 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
