

Roles of lymphatic endothelial cells expressing peripheral tissue antigens in CD4 T-cell tolerance induction
- PMID: 25857745
- PMCID: PMC4403767
- DOI: 10.1038/ncomms7771
Roles of lymphatic endothelial cells expressing peripheral tissue antigens in CD4 T-cell tolerance induction
Abstract
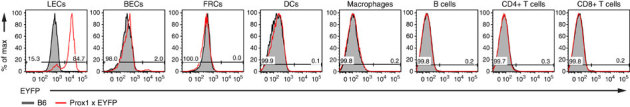
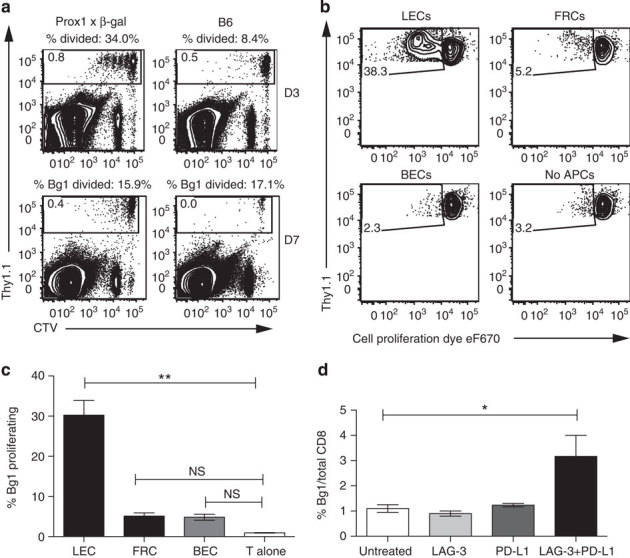
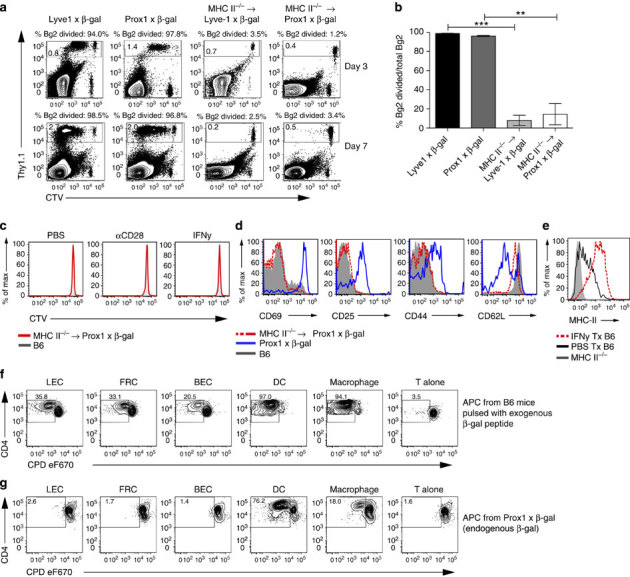
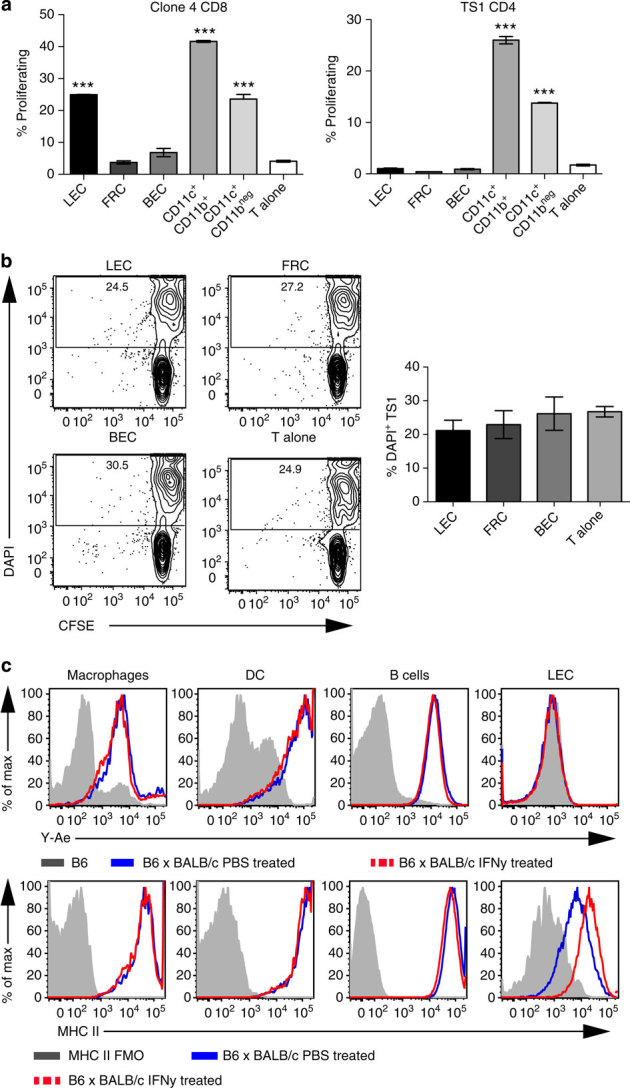
Lymphatic endothelial cells (LECs) directly express peripheral tissue antigens and induce CD8 T-cell deletional tolerance. LECs express MHC-II molecules, suggesting they might also tolerize CD4 T cells. We demonstrate that when β-galactosidase (β-gal) is expressed in LECs, β-gal-specific CD8 T cells undergo deletion via the PD-1/PD-L1 and LAG-3/MHC-II pathways. In contrast, LECs do not present endogenous β-gal in the context of MHC-II molecules to β-gal-specific CD4 T cells. Lack of presentation is independent of antigen localization, as membrane-bound haemagglutinin and I-Eα are also not presented by MHC-II molecules. LECs express invariant chain and cathepsin L, but not H2-M, suggesting that they cannot load endogenous antigenic peptides onto MHC-II molecules. Importantly, LECs transfer β-gal to dendritic cells, which subsequently present it to induce CD4 T-cell anergy. Therefore, LECs serve as an antigen reservoir for CD4 T-cell tolerance, and MHC-II molecules on LECs are used to induce CD8 T-cell tolerance via LAG-3.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Hinterberger M. et al. Autonomous role of medullary thymic epithelial cells in central CD4(+) T cell tolerance. Nat. Immunol. 11, 512–519 (2010) . - PubMed
-
- Ramsdell F. & Fowlkes B. J. Clonal deletion versus clonal anergy: the role of the thymus in inducing self tolerance. Science 248, 1342–1348 (1990) . - PubMed
-
- Bonasio R. et al. Clonal deletion of thymocytes by circulating dendritic cells homing to the thymus. Nat. Immunol. 7, 1092–1100 (2006) . - PubMed
-
- Baba T., Nakamoto Y. & Mukaida N. Crucial contribution of thymic Sirp alpha+ conventional dendritic cells to central tolerance against blood-borne antigens in a CCR2-dependent manner. J. Immunol. 183, 3053–3063 (2009) . - PubMed
-
- Derbinski J., Schulte A., Kyewski B. & Klein L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2, 1032–1039 (2001) . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 GM007267/GM/NIGMS NIH HHS/United States
- R21 AI109250/AI/NIAID NIH HHS/United States
- AI109250/AI/NIAID NIH HHS/United States
- T32 AI007496/AI/NIAID NIH HHS/United States
- 5 P30 CA044579/CA/NCI NIH HHS/United States
- CA009109/CA/NCI NIH HHS/United States
- AI068836/AI/NIAID NIH HHS/United States
- AI099033/AI/NIAID NIH HHS/United States
- P30 CA044579/CA/NCI NIH HHS/United States
- R01 AI068836/AI/NIAID NIH HHS/United States
- AI07496/AI/NIAID NIH HHS/United States
- Cancer Research UK/United Kingdom
- R56 AI099033/AI/NIAID NIH HHS/United States
- GM007267/GM/NIGMS NIH HHS/United States
- T32 CA009109/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
